Science 2024 | 靶点条件扩散生成高效TNFR超家族拮抗剂与激动剂
今天介绍的是Baker发表在Science (2024.10.6)的一项研究,展示了如何利用条件扩散模型成功生成针对TNFR超家族的高效拮抗剂与激动剂。TNFR家族成员在炎症和免疫调控中扮演关键角色,却因表面平坦、疏水性残基稀少而一直是蛋白设计中的“硬骨头”。传统方法往往依赖小型骨架和随机筛选,难以实现与天然复合物相媲美的界面匹配。该研究创新性地采用从随机残基云出发的自由扩散与部分扩散策略,在完全不依赖预设骨架的情况下生成大界面、高形状互补的结合物。结果显示,设计物在TNFR1上达到了皮摩尔级超高亲和力,并可通过部分扩散实现对TNFR2、OX40和4-1BB的特异性切换。当这些结合物进一步多价化时,能够分别表现为强效拮抗剂或超激动剂。更重要的是,该策略极大缩减了设计与筛选规模,却获得了远超既往成果的结合性能。这不仅为炎症与免疫疾病提供了全新治疗候选,还预示着蛋白设计正在进入以计算生成取代传统免疫与大规模筛选的新时代。

获取详情及资源:
0 摘要
尽管蛋白结合蛋白的设计取得了一定进展,但其与目标的形状匹配度仍低于许多天然蛋白复合物,尤其在肿瘤坏死因子受体1(TNFR1)及其他表面相对平坦且极性的蛋白靶点上,设计尝试多以失败告终。该研究提出假设:从随机噪声出发的自由扩散过程,或许能够为这类难缠靶点生成高度匹配的结合物,并以TNFR1为例进行了验证。结果显示,该方法获得了低皮摩尔亲和力的设计,并且通过部分扩散即可将其特异性完全切换至其他家族成员。当这些设计以更高价数形式呈现时,能够在OX40与4-1BB上分别表现为拮抗剂或超激动剂。这种能够在体外实现对药理学重要靶点的高亲和力与高特异性拮抗剂和激动剂的设计能力,预示着蛋白设计即将进入一个新时代——结合物将更多依赖计算生成,而非免疫或随机筛选。
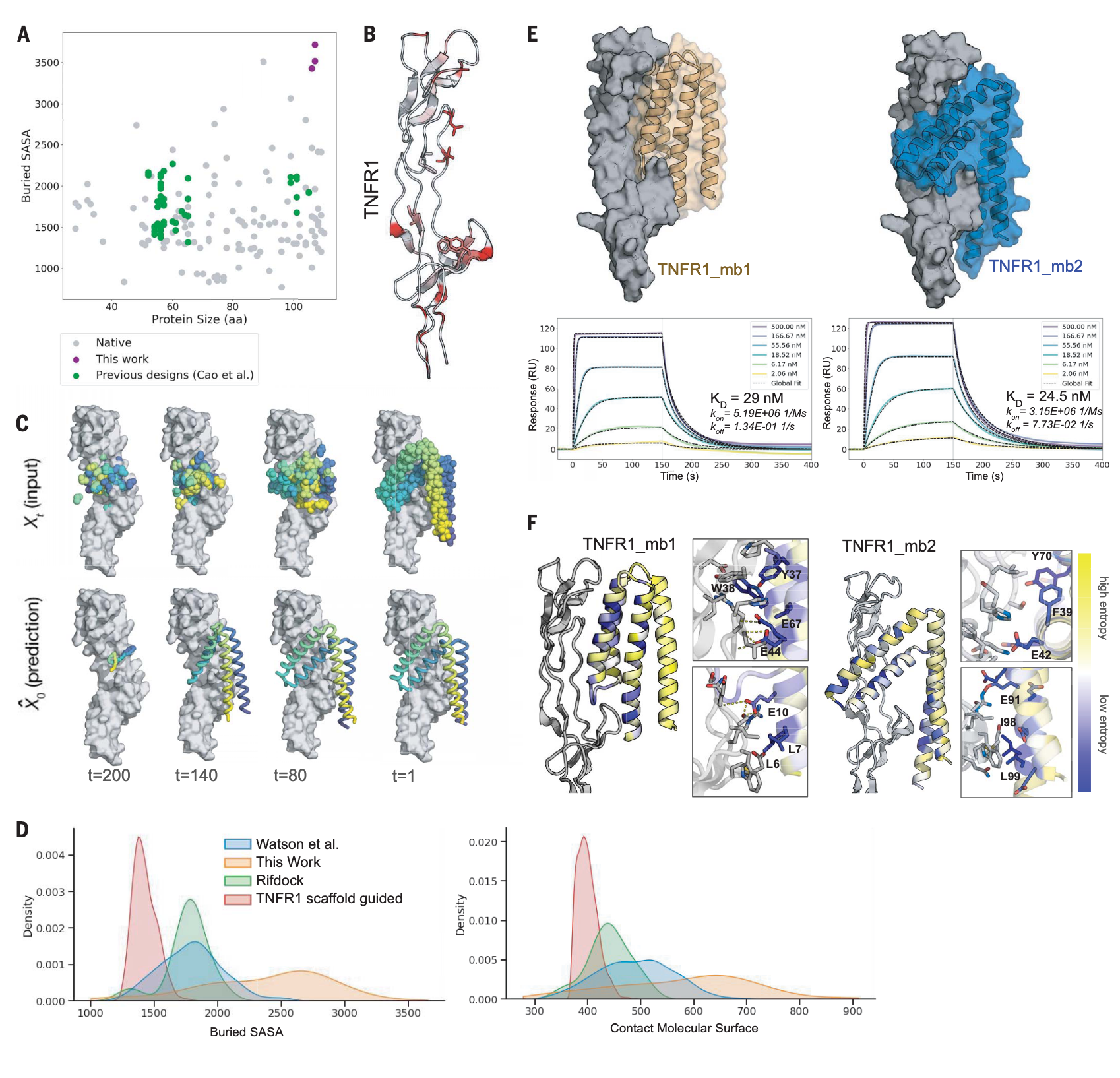
图1 | 形状互补结合蛋白的扩散过程 (A) 以往的 de novo 设计结合物(绿色)通常通过对预生成骨架进行对接产生,其与靶点埋藏的表面积(Ų)远小于许多天然复合物(灰色)。该研究的方法能够设计出界面更大的结合物(紫色)。(B) TNFR1是一个极具挑战性的靶点,其表面相对平坦,疏水残基很少(红色显示,PDB ID: 6KP8)。RFdiffusion所选定的热点残基以棒状表示。(C) 以TNFR1为靶点的代表性RFdiffusion轨迹。从随机残基分布开始(左上),在每一步去噪过程中(Xt, 上方),网络生成预测结构(^X0, 下方),并逐步插值趋近该结构以生成下一步(Xt-1)。(D) 与之前的TNFR1设计尝试比较。RFdiffusion生成的TNFR1结合物(橙色)在埋藏SASA与CMS指标上均明显高于使用Rosetta RIF dock方法生成的设计(红色),后者未能成功结合。图中还显示了RIFdock在多个靶点上的结果(绿色)以及短链RFdiffusion的结果(蓝色)。(E) 结合物TNFR1_mb1(橙色)与TNFR1_mb2(蓝色)的设计模型与TNFR1(灰色)复合物的结构示意(上)。表面等离子体共振(SPR)实验结果显示其与TNFR1的结合性能(下)。(F) 饱和突变(SSM)验证结合物设计模型与相关熵。TNFR1_mb1与TNFR1_mb2的所有单氨基酸突变体在酵母表面表达,并通过流式分选结合生物素化TNFR1,再进行深度测序。结果显示,核心及结合界面的残基高度保守(低熵,蓝色),而远离界面的多数表面残基则表现出高熵(黄色)。局部放大图展示了低熵残基形成的致密相互作用网络。
1 引言
高亲和力与高特异性结合蛋白的设计一直是计算结构生物学中的长期挑战,其应用范围涵盖治疗、诊断乃至更广泛的领域。传统的蛋白设计方法通常依赖于预先存在的骨架——包括天然蛋白或 de novo 设计的构件——这些骨架具有明确定义的三级结构。例如,早期的结合物设计是通过将大量理想化的约65个残基的蛋白骨架对接至靶点,并筛选出能够形成最低能量结合相互作用的对接构型。然而,这种依赖骨架的方法在应对表面平坦、缺乏凹槽的靶点时,往往受限于形状匹配能力。事实证明,迄今为止的 de novo 结合蛋白,其**接触分子表面积(CMS)**普遍低于许多天然复合物(图1A)。
新兴的生成式人工智能方法RFdiffusion为这一困境提供了新思路。它通过逐步去噪随机残基的空间分布来生成新的蛋白结构,并能结合条件信息引导,生成多样化的蛋白结构与功能。在设计结合物时,扩散轨迹会在目标蛋白固定存在的条件下运行,从而推动蛋白折叠出与靶点互补的构象。尽管此前RFdiffusion多依赖小型理想骨架进行条件生成,完全无约束扩散仅能得到简单的三或四螺旋束,但研究者推测,如果完全从随机残基分布出发,并在仅以目标结构作为约束条件的情况下进行扩散,或许能够生成更大、更复杂的折叠,甚至实现围绕平坦表面的大范围形状匹配。
因此,该研究尝试探索一种新的思路:不依赖预设骨架,也不受残基数量限制,仅以目标蛋白结构作为条件,直接利用RFdiffusion生成具备大界面、强形状匹配的折叠,从而实现针对以往设计失败的难点靶点的高亲和结合物设计。
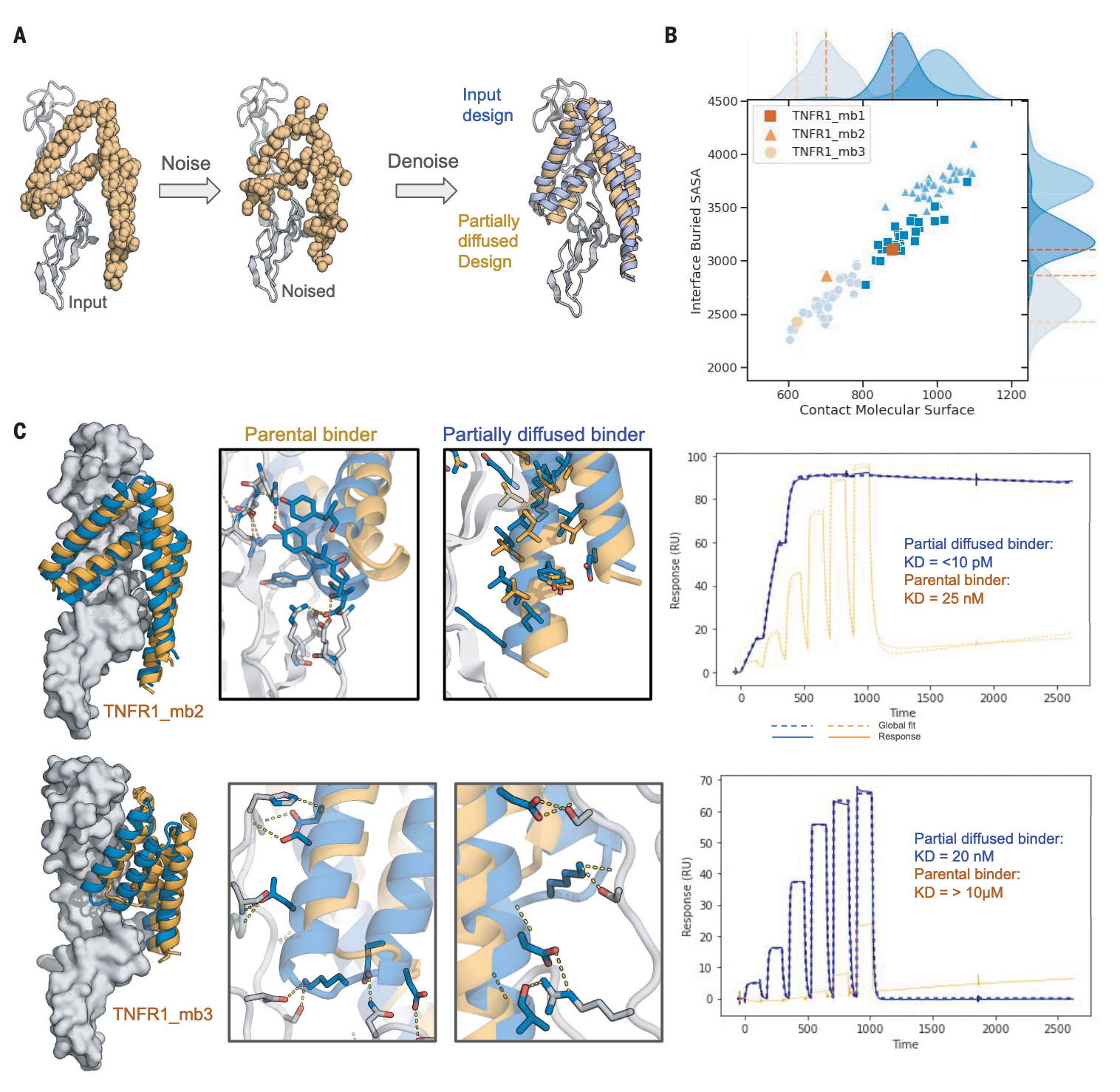
图2 | 部分扩散生成皮摩尔级结合物 (A) TNFR1_mb2的部分扩散示意。输入结构骨架表示为独立残基集合(左),加入噪声(中),经RFdiffusion去噪后得到与输入(紫色)相似但更契合靶点的新模型(橙色,右)。(B) 部分扩散增加了界面接触。输入设计(橙色)与部分扩散变体的界面接触分布显示在蓝色方块(TNFR1_mb1)、三角(TNFR1_mb2)、圆点(TNFR1_mb3)中。(C) 部分扩散增强了界面相互作用密度及结合亲和力。TNFR1_mb2形成了额外界面,而原有界面基本不变;TNFR1_mb3的形状匹配改善则带来额外作用。SPR实验结果表明亲和力显著提升。
2 结合物设计
研究重点放在肿瘤坏死因子受体超家族(TNFRSF),其中包括多个重要的药物靶点,尤其是TNFR1,其在炎症性疾病中发挥关键作用。与该家族其他成员类似,TNFR表面平坦且高度极性,缺乏疏水性凹槽,这使得以往的de novo设计方法难以奏效(图1B)。早前尝试利用RFdiffusion结合65个氨基酸骨架库或限制在小于65个残基的短链进行设计,但并未成功(fig. S1)。
为突破这一限制,研究者调整了RFdiffusion,使其能够生成具有更大接触面的骨架,以捕捉分散的表面疏水残基。蛋白相互作用主要受疏水作用驱动,而TNFR表面少数疏水残基在空间上相隔甚远,距离可达28 Å,这超出了65残基小型蛋白同时作用的能力。因此,在天然配体(TNF)界面处放置最多120个残基的高斯随机残基云,并通过RFdiffusion引导其与这些分散的疏水残基形成接触,从而生成与TNFR1形状高度互补的骨架(fig. S2)。随后,利用ProteinMPNN为这些骨架设计序列,以保证正确折叠并与靶点结合。
为了在有前景的设计附近进一步采样,研究者参考了 AlphaFold2(Af2)的复合物预测。若界面区域预测对齐误差(pae_interaction)低于20,则说明扩散过程在约200步中的第120步时结构已接近最终构象,于是从这些中间步骤提取骨架进行优化(图1C)。最终筛选出界面 pae<7.5 且 pLDDT>85 的设计用于实验验证(table S1),其接触分子表面积(CMS)与埋藏溶剂可及表面积(SASA) 均远高于以往设计(图1D)。
研究合成了96个设计的基因并在大肠杆菌中表达,其中90个表达良好且以单体为主(fig. S3A)。表面等离子体共振(SPR)实验显示,其中6个能够结合TNFR1(fig. S3B),其中TNFR1_mb1与TNFR1_mb2的解离常数(KD)分别为29 nM与24.5 nM,且对TNFR2无可检测结合,表现出高度特异性。与之前的de novo设计相比,这些结合物的三级结构迥异,并且与TNFR1形成了大范围界面作用(图1D)。值得注意的是,TNFR1_mb2呈现罕见的V形折叠,其CMS高达795 Ų,显著超过此前设计平均的490 Ų;TNFR1_mb1则在结合裂缝中插入一条环结构,CMS更高达897 Ų。
为了精确验证结合模式,研究对两个设计的每个残基进行了单点突变,总计4047个替换(figs. S4, S5)。结果与模型高度一致:影响结合的突变主要集中在设计界面及蛋白核心(核心突变会破坏折叠)。两种设计的关键相互作用均集中在TNFR1的107和111号疏水残基及38和40号残基,并辅以极性相互作用稳定界面(图1E, table S2)。
总体来看,饱和突变结果与设计模型界面高度吻合,且AF2与RoseTTAFold2的复合物预测进一步确认了结合模式(pAE分别为5.1与4.1;CA-RMSD分别为0.6 Å与1.0 Å)。这些结果强有力地表明,TNFR1_mb1与TNFR1_mb2确实按照设计模式与TNFR1结合。
2.1 计算机模拟的亲和力成熟
TNF-α以三聚体形式结合TNFR1,其亲和力高达19 pM。若要用单体蛋白在炎症中有效竞争这一结合,则需要更高的亲和力。为优化TNFR1_mb1、TNFR1_mb2和TNFR1_mb3的结合性能,研究并未依赖饱和突变(SSM)组合筛选,而是采用部分扩散策略(图2A)。通过在15–25步中向骨架加入噪声,再经RFdiffusion去噪,得到既类似又区别于原始设计的新骨架(RMSD为0.58–4.96 Å)。每个起始结构生成25,000个部分扩散骨架,经ProteinMPNN设计并由AF2预测界面pae_interaction <5的序列用于实验。结果显示,这些设计在CMS和埋藏SASA上均明显优于母体结构(图2B)。96个设计中94个在大肠杆菌中高水平表达,30%能与TNFR1结合。部分扩散使TNFR1_mb2的亲和力提升约三数量级,达到低于10 pM的水平,而TNFR1_mb3也由微摩尔级提升至20 nM(图2C)。对于TNFR1_mb2,这一改进源于额外界面形成及整体契合度的提高。部分扩散生成的TNFR1_mb2_pd1,成为迄今为止亲和力最高的单体TNFR1结合蛋白。
2.2 特异性切换
在亲和力优化成功的基础上,研究进一步探索是否能将结合特异性切换至TNFR家族其他成员。通过将TNFR1设计物叠合至TNFR2、OX40和4-1BB,并运行25,000条部分扩散轨迹,生成并筛选出AF2预测可靠的设计。在TNFR2中,32%的设计能高特异性结合,最高亲和力达198 pM,而对其他家族成员无作用。对于OX40,尽管天然配体界面差异显著,仍得到KD为30 nM的结合物,且其三级结构和结合角度相较母体设计发生更大调整。对于4-1BB,则需要额外的部分扩散轮次,但依然获得了22个高特异性结合设计,最高亲和力为44 nM。代表性设计4-1BB_mb1在骨架中引入弯折螺旋和短β折叠,以适应其独特的受体结构。
2.3 近原子分辨率的TNFR2设计
通过X射线晶体学解析了亲和力最高的TNFR2_mb1与TNFR2复合物的结构,结果与计算模型高度一致,结合物的C-α RMSD仅为0.52 Å,界面关键侧链相互作用与设计几乎完全重合(图3D)。
2.4 信号通路拮抗
凭借皮摩尔级亲和力,TNFR1结合物被测试用于抑制炎症信号。在HEK293报告细胞系中,这些单体结合物显著抑制TNF-α信号,最佳设计的IC50为106 pM(图4A)。其热稳定性与血清稳定性表现优良,95℃加热30分钟或在小鼠血清中孵育2小时后仍保持活性。同时,疏水作用色谱显示其表面疏水性并不显著,低于阿达木单抗、依那西普等临床抗体(fig. S8B)。此外,TNFR1_mb2_pd1在家族内展现高度特异性,且在浓度高达100 nM时,对TNFR1敲除细胞系也未表现明显非特异结合(fig. S8C)。
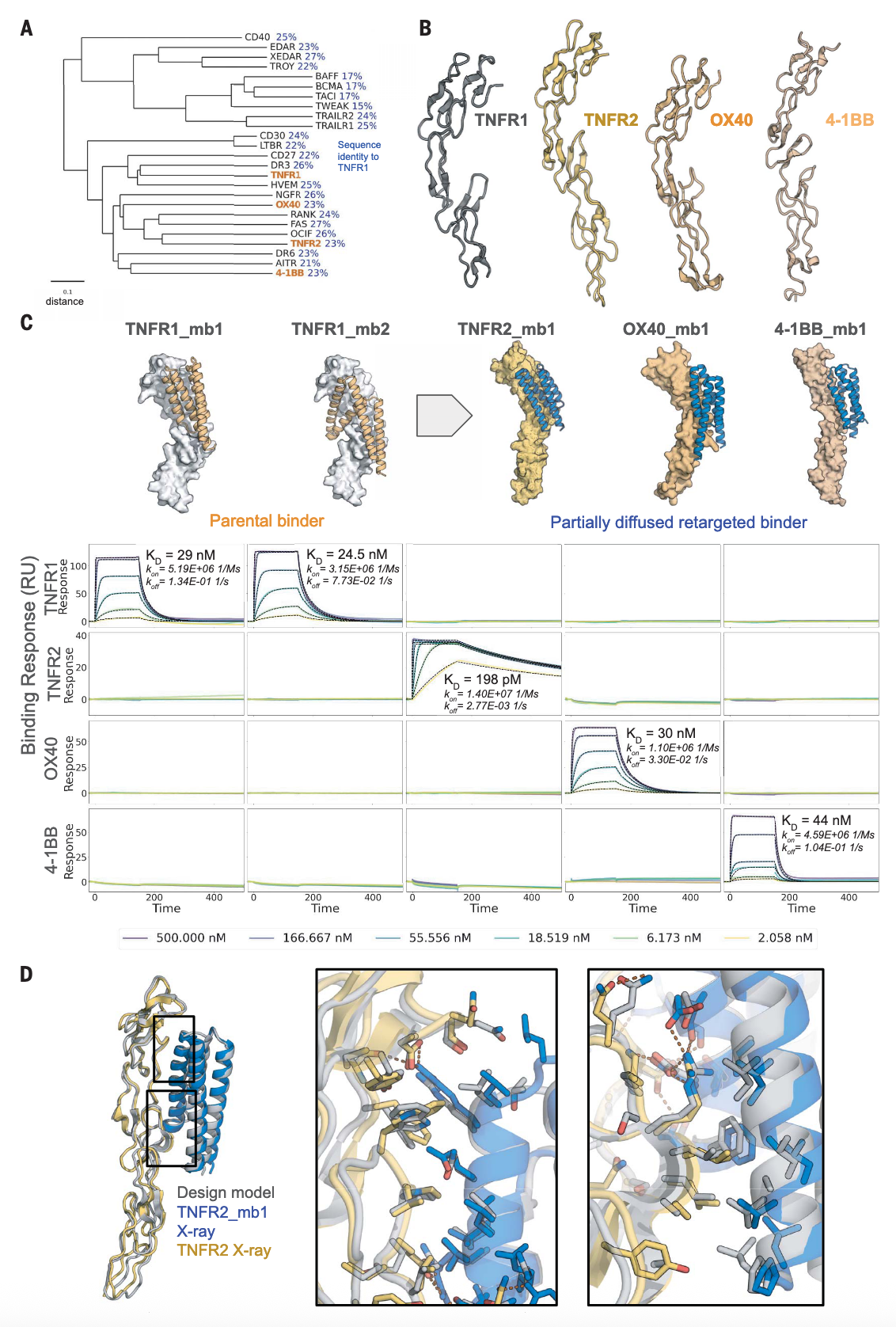
图3 | 部分扩散生成高特异性TNFR2、OX40与4-1BB结合物 (A) 基于ClustalW多序列比对与UPGMA法构建的TNFR超家族系统发育树。TNFR1、TNFR2、OX40和4-1BB以橙色标出。蓝色标注为与TNFR1的序列同一性。(B) TNFR1、TNFR2、OX40和4-1BB的结构比较(PDB IDs: 7KP8, 3ALQ, 2HEV, 6BWV)。(C) (上) 针对TNFR2、OX40和4-1BB的高亲和结合物原始设计模型与部分扩散后的模型。TNFR1的两个结合物(橙色)被叠加到新靶点上,经部分扩散后得到目标匹配的骨架(蓝色)。(下) SPR实验显示结合物对相应扩散靶点具有高度特异性。(D) TNFR2_mb1(蓝色)与TNFR2(黄色)复合物的晶体结构,与计算设计模型(灰色)叠合。放大图展示了界面残基的对齐情况。
2.5 激动剂设计
对于OX40和4-1BB这两类在肿瘤治疗中被广泛研究、用于扩增T细胞的受体,其激动剂具有潜在的治疗价值。与TNFR1不同,4-1BB信号并不能仅由可溶性的三聚配体激活,其生理性配体位于相邻细胞的质膜上,通过诱导4-1BB形成具有更长程有序排列的结构来实现信号传导(图4B)。已有研究利用抗体介导的配体网络来驱动更高阶复合物的形成从而实现信号传递,虽然这类配体是可溶性的,但其构象却相当异质。
为探索结构清晰、单分散的4-1BB激动剂,研究者将4-1BB结合模块与不同价态及不同间距的设计型同源寡聚体融合(图4F红点所示)。结果发现,单体以及C2或C3寡聚体均不能触发信号,这与天然三聚配体无活性的情况一致。相比之下,C4、C5、C6和C8寡聚体能够诱导信号(图4C与图S10A),其中C6构建体表现出最强信号,它在保持与天然配体相似的受体间距的同时,提供了更高的价态(六个拷贝而非三个,见图4E)。在44种寡聚体构建体的筛选中(图4F),价态是决定信号强度的关键因素:C1-C3均无信号,C4可产生稳定但较弱的信号,而C5与C6则显著增强。除了价态,结合几何构型同样影响信号传导效率。若高阶寡聚体将受体间距拉大到超过天然配体的范围,则信号显著减弱或消失;若构型可能与受体冲突或穿透膜结构,同样不能产生信号(图4F)。这一结果与细胞内六聚体构象推断的信号机制相一致(图4B),但仍需进一步解释为何仅增加一个亚基(C4情况)即可实现激动效应。
在OX40的实验中,情况则截然不同。与4-1BB相比,三聚构建体本身就是有效的激动剂,这与OX40能够被可溶性三聚配体激活的事实相符。单体与二聚结合模块无法触发信号,而含有三、四或五个结合模块的寡聚构建体均能高效激活信号(图4D、4F及图S10B)。
无论是4-1BB还是OX40,其EC50(半数有效浓度)与Emax(最大信号强度)在不同寡聚构建体之间差异显著,这为通过调节价态与几何构型来精细调控信号反应提供了重要机会,不仅有助于深入理解这一重要受体家族的作用机制,也对治疗应用具有价值。尤其值得注意的是,最佳OX40和4-1BB合成激动剂的Emax远高于天然配体(OX40情况)或抗体-配体组装体(4-1BB情况),这类分子在T细胞扩增方面展现出特别有前景的应用潜力。
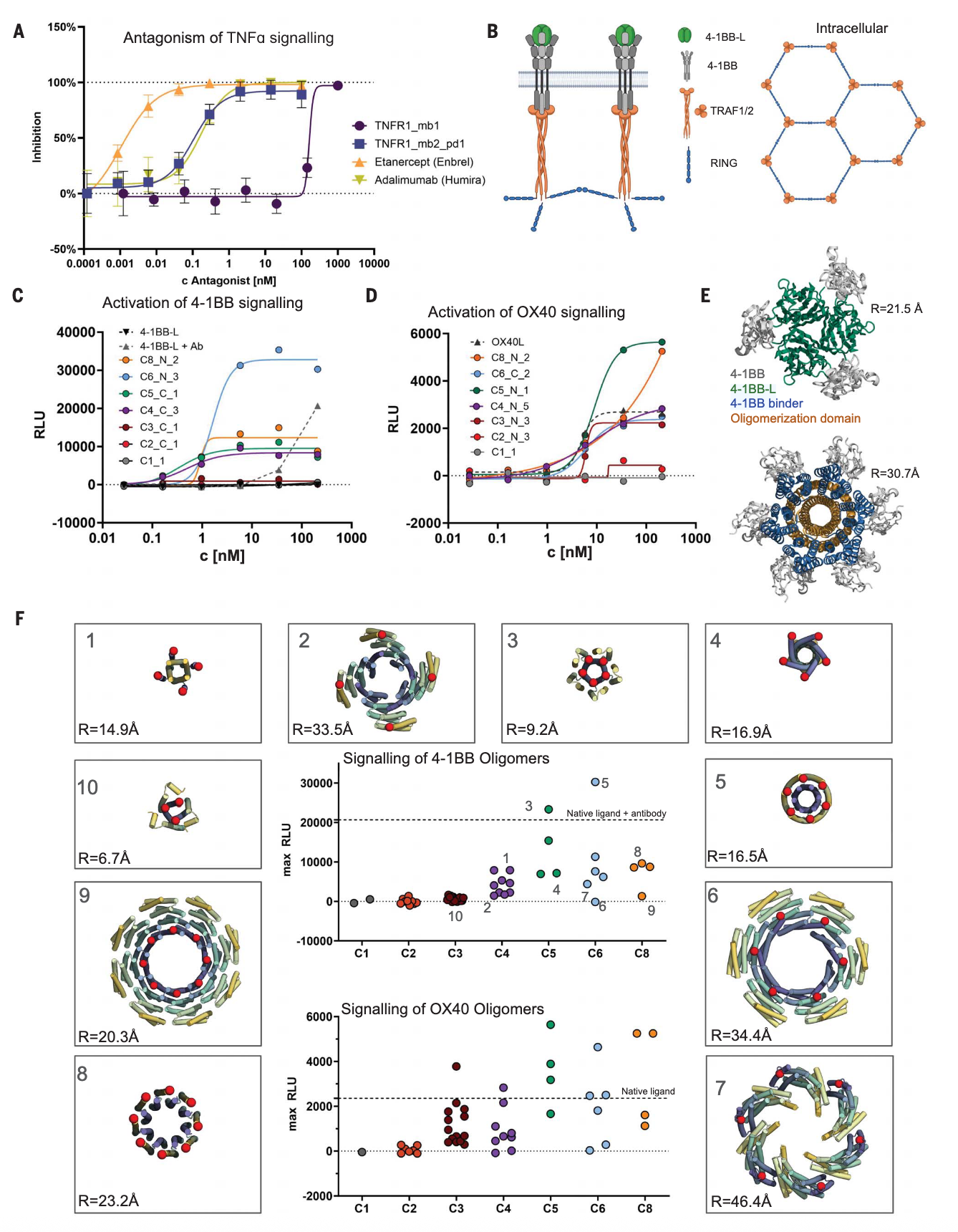
图4 | 可溶性寡聚体4-1BB与OX40超激动剂的设计 (A) 设计的结合物能够拮抗TNF-α信号。在HEK293-blue细胞中加入100 pM TNF-α,并测试不同浓度结合物对NF-κB依赖激活的抑制效果。(B) 4-1BB信号通路示意。三聚体配体(绿色)结合三个受体(灰色),诱导TRAF1/2三聚体(橙色)及锌-RING指结构六聚体形成,从而启动下游信号。(C) 将4-1BB结合物多价呈现于环状寡聚体上,可激活荧光素酶报告细胞系的4-1BB信号。设计物4-1BB_mb2融合不同的寡聚化结构域,价数从1到8不等。(D) OX40结合物多价呈现后,同样激活OX40信号。OX40_mb1融合不同寡聚化结构域进行测试。(E) 模型显示,4-1BB_mb2(蓝色)融合至环状六聚体(橙色)N端,与4-1BB(灰色)结合后,与天然配体复合物(绿色, PDB ID: 6BWV)相比,结合构象显著不同。(F) 设计的4-1BB与OX40激动剂表现出几何结构与寡聚化状态依赖性。在200 pM条件下的最大信号显示,不同寡聚化状态对活性影响显著。中心图以点状分布表示不同融合物的信号强度,并与天然配体进行比较,周围面板展示了寡聚化结构域的模型示例。
3 讨论
在某些治疗挑战中,若必须实现拮抗而完全避免激动风险,高亲和力的单体结合物相较于二价抗体具有优势,因为后者可能二聚化靶受体并激活信号。TNF-α及其受体TNFR在炎症性疾病中发挥核心作用,因此是重要的药物靶点。现有疗法主要选择作用于配体TNF-α而非TNFR1,以避免潜在的炎症反应激活。然而,这种方式同时也抑制了通过TNFR2介导的潜在抗炎信号,可能导致该类药物产生不良副作用。
该研究中的TNFR1结合物具有非常大的界面,甚至超过了天然复合物的结合界面,尽管其长度仅为107个氨基酸。这种特性可能使其实现飞摩尔(fM)级别的超高亲和力拮抗。此外,这类小型设计蛋白具备高稳定性和可能的低生产成本,有望实现口服递送,在肠道疾病治疗中具有应用前景。
正如4-1BB与OX40超激动剂所展示的那样,高亲和力单体结合物可用于构建多种可溶性信号分子,相较于目前依赖天然配体与交联抗体的策略,这种方法提供了更强的可控性。更广泛地看,能够在无需免疫动物、筛选大规模随机文库或测试成千上万设计候选的情况下,生成对治疗相关且结构复杂靶点具有高亲和力和高特异性的结合物,标志着结合物设计与治疗性候选分子发现进入了新纪元。
在实验规模方面,该研究测试的序列数量远少于以往:TNFR1的96个自由扩散设计与96个部分扩散设计,以及TNFR2、OX40与4-1BB的48个部分扩散设计,相较于过去依赖展示体系筛选上万设计的策略,规模显著缩小,且未进行随机或实验指导下的优化,仅通过在第一轮96个设计中选择最佳者进入部分扩散步骤。
该研究获得的TNFR1 <10 pM与TNFR2 198 pM的亲和力,是迄今报道的单体结合物在这些靶点上的最高水平。相比之下,可能以二价方式结合的抗体亲和力可达680 pM,而单体形式的Fab、scFv和纳米抗体仅在10~100 nM范围。这种超高亲和力,很可能来源于极高的形状互补性与埋藏表面积。事实上,这些设计在靶点上埋藏的表面积远高于以往的minibinder设计,且每个残基的埋藏面积已可与天然复合物媲美,而后者经历了数亿年的进化(图1A)。
结合从随机残基云(>100个氨基酸)出发的RFdiffusion与部分扩散来优化亲和力和家族成员特异性,构成了一种极其强大且通用的策略,能够获得针对结构挑战性靶点的高效能、高亲和力结合物。