Nat. Microbiol. 2025 | APEX: 深度学习揭示古菌蛋白组中的抗生素
今天介绍的是发表在 Nature Microbiology 的一项研究,研究团队利用深度学习框架 APEX 1.1 系统性地探索了古菌蛋白质组,首次揭示古菌中存在大量此前未被识别的抗菌分子。与传统主要依赖细菌和真菌的抗生素来源不同,古菌因其独特的进化地位和生化特征,长期以来几乎未被开发。该研究从233个古菌蛋白组中预测出超过12,000条潜在抗菌肽,并合成了其中80条进行实验验证,结果显示其中93%在体外表现出显著抗菌活性,可有效抑制多种临床耐药病原体。更为重要的是,在小鼠感染模型中,部分候选肽如 archaeasin-73 能够显著降低鲍曼不动杆菌的负荷,其疗效可与多黏菌素B等临床常用抗生素相当,同时未观察到明显毒性。这一成果凸显了古菌作为新一代抗生素重要来源的巨大潜力,也展示了深度学习结合实验验证在加速抗菌药物发现中的突破性价值。

获取详情及资源:
- 论文: https://doi.org/10.1038/s41564-025-02061-0
- 代码: https://gitlab.com/machine-biologygroup-public/apex-pathogen
0 摘要
抗菌素耐药性是当今人类面临的最严重威胁之一,因此开发新型抗生素的需求愈发迫切。虽然大多数抗生素来源于细菌和真菌,但古菌却提供了一个几乎未被开发的抗生素资源。在该研究中,研究者利用深度学习系统性地探索了古菌组,挖掘出具有潜力的候选分子用于对抗抗菌素耐药性。通过对233个古菌蛋白组的挖掘,共鉴定出12,623种具有潜在抗菌活性的分子。这些被称为archaeasin的肽类化合物,展现出区别于传统抗菌肽的独特组成特征,包括显著不同的氨基酸构成。
研究者合成了其中的80种archaeasin,其中93%在体外实验中表现出抗菌活性,可作用于鲍曼不动杆菌、大肠杆菌、肺炎克雷伯菌、铜绿假单胞菌、金黄色葡萄球菌和肠球菌等病原体。值得注意的是,体内验证结果显示,archaeasin-73成为核心候选分子,在小鼠感染模型中能够显著降低鲍曼不动杆菌负荷,其疗效可与多黏菌素B等已知抗生素相媲美。
这一发现凸显了古菌作为开发新一代抗生素重要资源的巨大潜力。
1 引言
抗菌素耐药性的上升趋势已成为全球最紧迫的公共卫生威胁之一。耐药病原体削弱了现有抗生素的疗效,导致感染治疗难度不断增加。这一日益严峻的危机凸显了新型抗生素开发的关键需求。然而,在过去的几十年里,抗生素的发现进程显著放缓,传统的研发途径主要依赖于细菌和真菌。近年来,计算方法,尤其是深度学习模型,开辟了抗生素发现的新方向,使得研究者能够系统性地探索庞大的序列空间。
尽管古菌在进化中的重要性和生化多样性早已得到认可,但在抗生素发现领域仍然鲜有探索。与细菌和真核生物不同,古菌拥有独特的脂质膜、代谢途径和应激适应机制,这些特征可能会影响其加密肽(EPs)的结构和功能。由于独特的结构与功能特性,加密肽已成为抗生素研发的前沿热点。这类肽通常在常规的序列检索中被忽视,但在被正确识别和验证后却可能展现广谱的抗菌活性。已有研究表明,来源于人类、细菌甚至灭绝生物蛋白组的加密肽,都可以作为有效的抗菌分子。然而,目前尚未有系统性研究探索古菌组中的此类生物活性序列。基于古菌与细菌及真核生物在进化上的高度分化,其加密肽很可能具有区别于已知抗菌分子的独特结构与作用机制。
研究团队应用更新后的深度学习框架APEX 1.1,系统性地挖掘了来源于Swiss-Prot数据库的所有古菌蛋白组。通过这一方法,研究者识别出具有潜在抗菌活性的加密肽,并将其命名为archaeasin。利用基于已知抗菌肽(AMPs)与加密肽序列训练的计算管线,共从233个古菌蛋白组中预测得到12,623个潜在AMPs。在这些候选分子中,研究者合成并实验测试了80种archaeasin,其中93%在体外实验中展现抗菌活性。进一步的体内验证表明,archaeasin-73在小鼠感染模型中能够有效降低病原菌负荷,其抗菌效果与多黏菌素B相当。
通过将加密AMPs的探索扩展到古菌组,该研究不仅揭示了这一生命领域尚未开发的巨大潜力,也展示了深度学习与实验验证结合加速抗生素发现的强大能力。这些结果提供了大规模证据,证明古菌编码了丰富的具有潜在治疗价值的肽类库,凸显了其作为新一代抗生素来源的重要意义。
2 结果
2.1 深度学习指导下的archaeasin鉴定
研究团队从UniProt数据库中收集了来自233种古菌的18,677条非冗余已审查蛋白序列,并利用更新后的深度学习抗菌活性预测器APEX 1.1(Methods中称为“APEX 1.1”)对古菌蛋白组中的加密肽(EPs)进行挖掘。由于APEX能够预测细菌株特异的最低抑菌浓度(MIC),因此研究者采用平均MIC来代表肽类的整体抗菌效力,并最终发现12,623条平均MIC≤100 μmol l−1的EPs。这些序列在被扫描的193,331,608条古菌肽中占比为0.00653%。为了评估该信号是否超出随机预期,研究者生成了一个长度分布与archaeasin相匹配的非冗余随机肽集,共193,288,387条。按照同样的AMP判定标准(平均MIC≤100 μmol l−1),从中仅鉴定出5,292条预测活性肽,占比0.00274%,约比古菌EP中低2.38倍。这些结果表明,古菌蛋白组中的抗菌序列相较随机抽样具有显著富集。
接着,研究者考察了基因组较大的古菌是否更可能编码更多预测的AMP。通过整合预测活性EP的数量(定义为在11种病原株中平均MIC≤100 μmol l−1的肽)与NCBI注释的蛋白编码基因总数,对预测EP数量最高与最低的10个古菌属进行了分析。Spearman相关性分析结果显示,基因组大小与预测EP数量之间存在显著正相关(ρ=0.4475, P=0.0479),提示基因组更大的古菌可能蕴含更丰富的潜在抗菌序列。值得注意的是,一些高评分的属如Pyrococcus、Methanocaldococcus、Pyrobaculum和Sulfolobus均为嗜热生物,这一现象暗示某些生活方式特征,如适应高温环境,可能与更高丰度的活性肽相关。尽管分析主要集中在基因组大小,但研究者也指出,环境因素如耐温性、代谢特化或生态位同样可能影响加密AMP的多样性与分布。这一结论仍属初步判断,随着更多古菌基因组被测序和注释,这些趋势将更加稳健,并有助于深入探索活性肽丰度的生态与进化相关性。
为了研究AMP样EP在古菌蛋白组中的分布,研究者首先对12,623条EP与来自DBAASP、APD3和DRAMP的19,775条公开AMP进行序列比对,并应用UMAP降维以可视化序列相似性矩阵。聚焦于前265条平均MIC<80 μmol l−1且序列多样性较高的archaeasin,研究者检索其来源蛋白的Gene Ontology注释。分析显示,这些高排名的archaeasin大多来源于细胞质蛋白及具有关键细胞功能的蛋白,包括ATP结合、金属离子结合、DNA结合、tRNA结合与锌离子结合。此外,还有一部分archaeasin来源于结构性核糖体蛋白、质膜蛋白以及参与翻译过程的蛋白。整体来看,结果突显了archaeasin在古菌细胞中分布广泛且功能多样。
在氨基酸组成方面,archaeasin展现出区别于数据库中已知AMP以及先前在人类蛋白组中发现的EPs的特征。它们显著富含谷氨酸残基,超过了通常AMP中的水平。这种高比例的负电荷残基在与人类EPs比较时同样出现。然而,archaeasin依然保留了阳离子残基的优势,使其整体上呈现比数据库AMP更高的正电荷比例,提示其在电荷分布上具有独特平衡。尽管存在这些差异,archaeasin的疏水性仍与标准数据库序列相当。此外,它们还表现出更强的两亲性倾向,即疏水与亲水残基之间分布更加均衡。
综上所述,该分析支持了这样一种观点:与人类类似,古菌也编码了丰富且组成独特的抗菌EP库。这一发现不仅揭示了古菌作为异常抗生素来源的潜力,同时也提供了跨越生命域理解抗菌多样性的进化学对照。
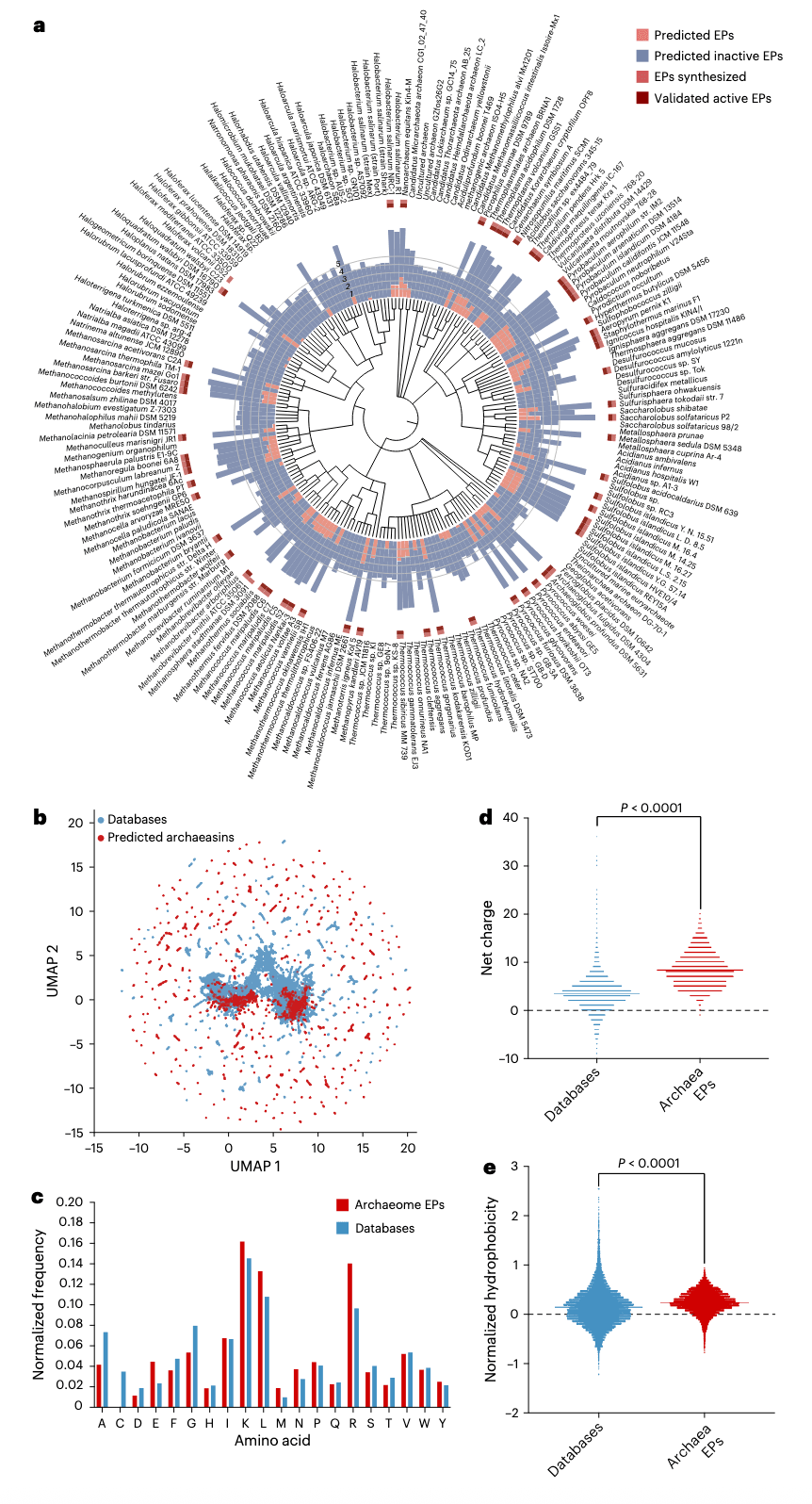
图1 | APEX对古菌组的探索 a,系统性扫描古菌蛋白质组以识别具有潜在抗菌活性的EPs。环形柱状图表示APEX发现的活性(红色)和非活性(蓝色)EPs的对数10转化平均值。若某肽对测试细菌株的预测平均MIC ≤100 μmol l−1,则被判定为活性。数值已根据每个被扫描生物的蛋白数量进行归一化。合成肽来源的古菌以浅红色方块标记,实验验证为活性的则以深红色方块突出显示。b,基于相似性矩阵的序列空间探索。图中展示了DBAASP数据库中的肽序列与APEX在古菌中发现的抗菌EPs的二维序列空间可视化。通过序列比对生成所有DBAASP肽与APEX预测的12,623条抗菌EPs的相似性矩阵(补充数据1与2)。矩阵中的每一行表示基于氨基酸组成的肽特征。随后应用UMAP降维至二维进行可视化(扩展数据图2a)。c,比较古菌EPs与已知AMPs(DBAASP、APD3和DRAMP 3.0数据库)的氨基酸频率(扩展数据图2b–e)。d, e,比较预测具有抗菌活性的肽与已知AMPs在两种理化性质上的分布:净电荷(d)与归一化疏水性(e)。净电荷影响肽与带负电的细菌膜之间的初始静电作用,而疏水性则影响其与膜双层脂质的相互作用(扩展数据图2)。氨基酸组成的差异通过卡方检验评估,P值为0,低于机器精度水平,提示差异具有统计学意义。d与e中的统计学显著性通过双尾t检验及Mann–Whitney检验确定,均为P < 0.0001。箱线图中实线表示各组的平均值。

图2 | 古菌来源抗生素的抗菌活性与二级结构特征 a,热图展示了古菌来源的活性抗菌剂对11种临床相关病原体的抑菌活性(μmol l−1),包括耐药的革兰氏阴性菌(标记为–)与革兰氏阳性菌(标记为+),其中包括耐碳青霉烯大肠杆菌(CRE)、耐甲氧西林金黄色葡萄球菌(MRSA)和耐万古霉素肠球菌(VRE)。实验中,将
2.2 archaeasin对细菌病原体的抗菌活性
为了在实验层面验证古菌EPs的抗菌活性,研究团队筛选了80条序列多样性较高(彼此序列相似度<70%)且在APEX 1.1中排名靠前的肽(Supplementary Data 2)。在化学合成与实验验证时,优先选择与已知AMP序列相似度低于70%的肽(Extended Data Fig. 2a)。此外,当两条挖掘出的序列相似性较高时,仅保留预测抗菌活性更强的一条肽(见Methods中的“Archaeasin selection”)。
这些archaeasin随后被用于针对临床相关病原体的体外测试,包括鲍曼不动杆菌、大肠杆菌、肺炎克雷伯菌、铜绿假单胞菌、金黄色葡萄球菌、屎肠球菌与粪肠球菌,浓度范围为1 μmol l−1至64 μmol l−1。结果显示,80条肽中有75条表现出抗菌活性(MIC ≤64 μmol l−1),至少能抑制一种病原菌株(Fig. 2a),命中率超过93%。多黏菌素B与左氧氟沙星被用作阳性对照(Extended Data Fig. 4a)。
此外,预测MIC与实验验证MIC之间的Pearson相关系数为0.503,证明了APEX 1.1的预测能力(Extended Data Fig. 4b)。进一步比较APEX首版与APEX 1.1在80条合成archaeasin上的实验与预测MIC之间的Pearson与Spearman相关性时,结果显示APEX 1.1的表现显著优于初版APEX(Supplementary Tables 3和4)。
这一部分实验验证明确支持了APEX 1.1在挖掘古菌抗菌肽方面的高效性与可靠性。
2.3 Archaeasin的无序与富含β构象的二级结构特征
短肽的二级结构往往具有动态性,会在疏水–亲水界面间转变于无序与有序构象,这种结构转变对肽的抗菌活性及其他生物学功能具有关键作用。为评估合成的archaeasin的二级结构,该研究在不同环境下进行了圆二色谱实验:水、含10 mmol l−1十二烷基硫酸钠(SDS)的水溶液,以及三氟乙醇(TFE)与水3:2 (v/v)的混合溶液。SDS胶束因与生物膜双层相似,被选作膜模拟环境;而TFE–水混合液则可通过脱水肽主链酰胺基团促进分子内氢键形成,从而诱导α螺旋结构。所有archaeasin均在50 μmol l−1浓度下,在190–260 nm波长范围内进行测试(扩展数据图5)。
二级构象比例的计算采用BeStSel服务器(Fig. 2b–d)。如预期,由于archaeasin序列较短(<50个氨基酸残基),在水中均呈无序状态(Fig. 2b);在其他两种环境中(即TFE–水混合液和SDS胶束),则呈现一定的β样构象倾向(fβ在20%–45%之间,Fig. 2c, d)。这种行为是典型的具有抗菌活性的短肽特征。与此前报道EPs主要表现为β样结构不同,archaeasin在诱导螺旋的环境及与脂质双层相互作用时则呈现α螺旋构象。
2.4 Archaeasin的功能协同与合作性作用
为了探究来自同一古菌株或其近缘菌株的分子是否能够协同增强彼此对病原体的抗菌活性,该研究进行了棋盘式实验。棋盘式实验是一种标准方法,通过在一系列浓度范围内测试两种抗菌剂的联合效应,以评估其相互作用。该方法能够定量判断组合是否表现出协同作用、相加效应、无关效应或拮抗效应。实验中肽的浓度范围从2倍MIC至32倍稀释,在与抗菌实验相同的条件下进行。
首先选择了鲍曼不动杆菌(A. baumannii)ATCC 19606株作为测试对象,该菌株以高度耐药性和作为全球重要的机会致病菌而闻名,并导致严重的院内感染死亡率。该菌株对archaeasin表现出高度敏感性。随后,根据系统发育树中亲缘关系较近(成对距离≤8)的原则,共选择了79对archaeasin进行检测(图3)。结果显示,大多数组合表现出协同或相加效应,由**分数抑菌浓度指数(FICI)**确定。FICI的常用分类为:FICI ≤ 0.5 表示强协同;0.5 < FICI ≤ 1 表示相加效应;1 < FICI ≤ 2 表示无关效应;FICI > 2 表示拮抗。FICI值越低,说明肽的联合效应越强,能显著提升抗菌效力。协同作用在抗菌研究中尤为重要,因为它既能降低单药用量、减少潜在毒性,又能延缓耐药发展、改善治疗效果。
值得注意的是,Methanocaldococcus属的archaeasin展现出最低的FICI值,如archaeasin-25与-27的组合FICI为0.25,archaeasin-21与-27、-28或archaeasin-24与-28的组合FICI均为0.375。类似地,Methanothermobacter属的组合FICI值在0.28至0.375之间;Thermococcus属的archaeasin-75与-76组合FICI为0.28;Pyrococcus属的archaeasin-60与-64、-65,archaeasin-61与-65,以及archaeasin-63与-64的组合FICI均为0.375。
结果显示,某些古菌谱系更容易产生协同肽。尤其是来自嗜热古菌(如Methanocaldococcus与Thermococcus)的肽,表现出最一致的协同作用。这提示适应极端环境的生物可能进化出了依赖合作机制的抗菌策略,以对抗竞争性的微生物群落。这些肽表现出协同效应的倾向,可能与其特殊结构适应性、在极端条件下的稳定性增强,或促进合作活性的理化性质相关。
这些发现强调了基于archaeasin的联合疗法在多重耐药感染治疗中的潜力,或能为抗菌药物开发提供新的替代方案。
2.5 Archaeasin的膜破坏作用机制
为了理解archaeasin对细菌细胞的作用方式,该研究通过荧光实验评估其是否涉及膜靶向机制。首先,在对鲍曼不动杆菌ATCC 19606的测试中,共鉴定出70条具有抗菌活性的archaeasin(图2a)。随后,检测这些肽在其MIC浓度下对细菌外膜与胞质膜的通透性(Fig. 4a,扩展数据图6a)与去极化作用(Fig. 4b,扩展数据图6b)。
为评估archaeasin对革兰氏阴性菌外膜的通透作用,采用N-苯基-1-萘胺(NPN)实验。NPN是一种疏水性染料,在富含脂质的环境(如细菌外膜)中会增强荧光。当外膜受损时,NPN能够进入并导致荧光增强(Fig. 4a)。结果显示,仅来自Methanocaldococcus jannaschii的archaeasin-21(母蛋白:未表征蛋白MJ1489.1)与archaeasin-22(母蛋白:推测为鞭毛相关蛋白D)能显著破坏细菌外膜。多黏菌素B作为阳性对照。总体来看,archaeasin的外膜通透能力弱于典型AMPs或人/动物来源的EPs。
随后采用3,3′-二丙基噻二羰氰碘化物(DiSC3-5)实验评估胞质膜去极化。当跨膜电位被破坏时,荧光探针迁移至胞外并导致荧光增强。在测试的70条肽中,有34条archaeasin使胞质膜去极化程度超过多黏菌素B对照组(Fig. 4b)。其中,来自Thermofilum pendens的archaeasin-78(母蛋白:丙氨酸tRNA连接酶)与来自Pyrobaculum arsenaticum的archaeasin-57(母蛋白:小核糖体亚基蛋白uS4)去极化效应最为显著。
进一步通过Laurdan广义极化实验支持了其膜靶向机制。结果显示,archaeasin处理后,鲍曼不动杆菌胞质膜的物理状态发生变化(Fig. 4c)。Laurdan荧光位移反映膜脂质堆积的变化,广义极化值增加提示膜刚性增强。与去极化数据一致,archaeasin-78与-57随时间推移显著提高广义极化值,说明这些肽不仅破坏膜电位,还直接扰乱膜脂质的有序性。相比之下,archaeasin-21与-22虽然能有效破坏外膜,但对胞质膜物理状态的影响有限,甚至表现出轻微下降。
这些差异与肽的结构特征及膜插入深度密切相关。结果表明,对胞质膜的相互作用(去极化与扰乱脂质组织),而非外膜通透性,是多数archaeasin的主要抗菌机制。这种作用方式类似于近期报道的小开放阅读框编码肽( smORF-encoded peptides ),而不同于传统AMPs与EPs通常靶向外膜的机制。
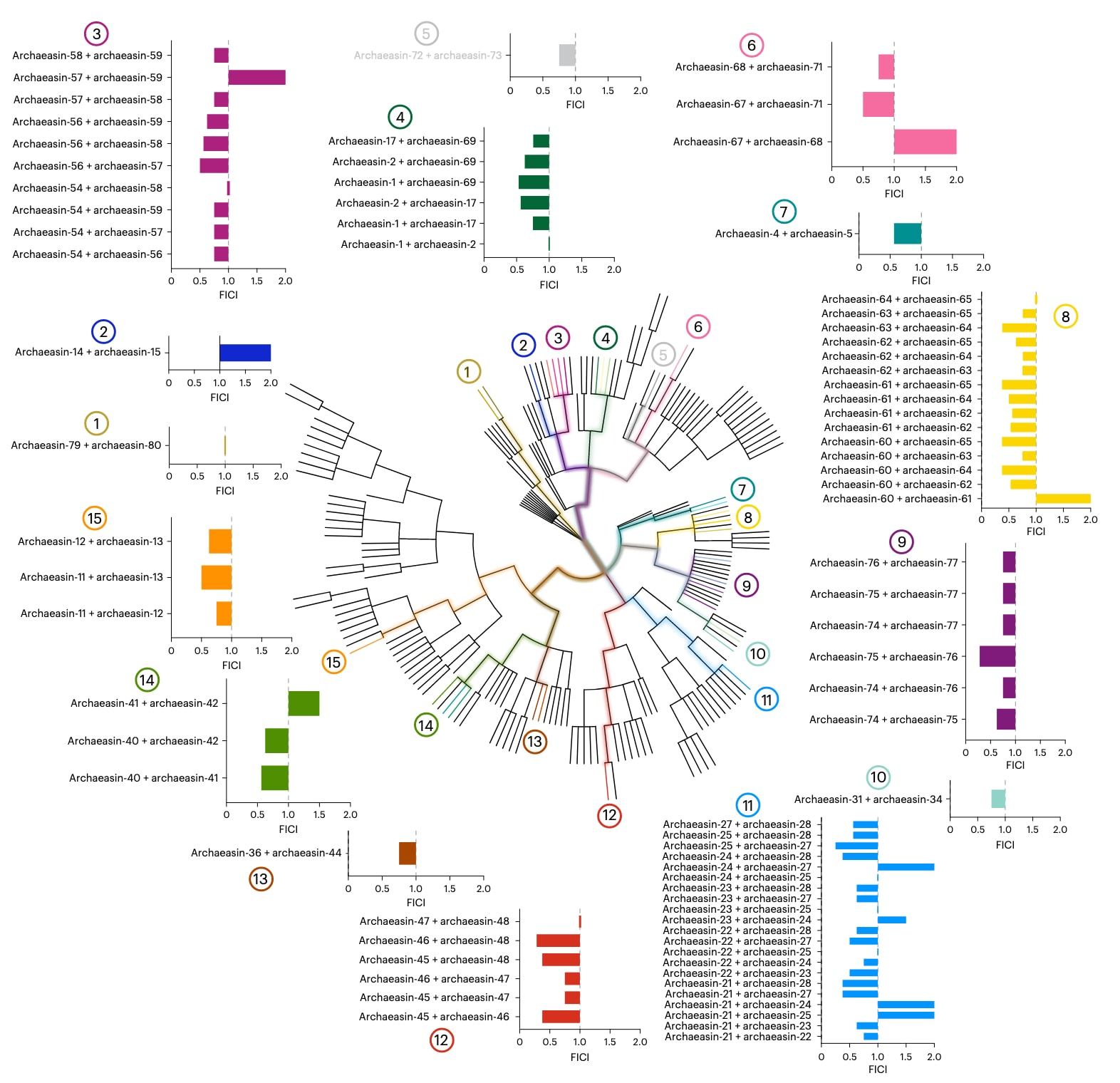
图3 | 古菌肽类抗生素之间的协同作用 通过棋盘式稀释实验评估来自同一或系统发育上密切相关的古菌(系统发育成对距离 ≤ 8)的肽类抗生素(EPs)对 鲍曼不动杆菌 ATCC 19606 的活性。实验采用两倍梯度稀释,从
2.6 古菌素在人类细胞系中的低毒性
为评估合成古菌素(archaeasins)的潜在毒性,首先采用人类红细胞(RBC)进行测试,这是评估抗菌剂毒性的常用方法。结果显示,在80种被检测的古菌素中,有25种(31.3%)在实验浓度范围内表现为中度至低水平的溶血活性,其 HC50 值(导致50% RBC溶血的肽浓度,经线性回归计算)均 ≤64 μmol l⁻¹(见图4d与扩展数据图7)。值得注意的是,多数在低 MIC 值下对细菌病原体表现活性的古菌素,并未在相应浓度下显示明显的毒性作用(扩展数据图7)。然而,7种古菌素(archaeasin-12、-13、-32、-54、-58、-64 和 -78)确实表现出溶血效应。
为了进一步评估其安全性,又对古菌素在人类胚胎肾细胞(HEK293T)上的细胞毒性进行了检测。通过测定 CC50 值(使 HEK293T 细胞活性降低50%的肽浓度),获得了80种古菌素的毒性数据(见图4d)。这些数据以热图形式总结,标示出在何种浓度范围内观察到细胞毒性效应(图4d与扩展数据图7)。结果显示,大多数古菌素表现出低细胞毒性,CC50 值超过128 μmol l⁻¹,其中26种古菌素的CC50值高于128 μmol l⁻¹,表明在测试浓度范围内几乎不具备显著毒性。
然而,部分古菌素表现出低至中度的细胞毒性。尤其是与溶血效应重合的那7种古菌素(archaeasin-12、-13、-32、-54、-58、-64 和 -78),其 CC50 值均 ≤64 μmol l⁻¹,提示可能存在非靶向毒性。值得注意的是,多数具备强抗菌活性的古菌素(低 MIC 值)在相应浓度下对 HEK293T 细胞并未表现出明显毒性。这种选择性活性凸显了它们作为候选抗菌剂的潜力,同时对人类细胞的副作用有限。
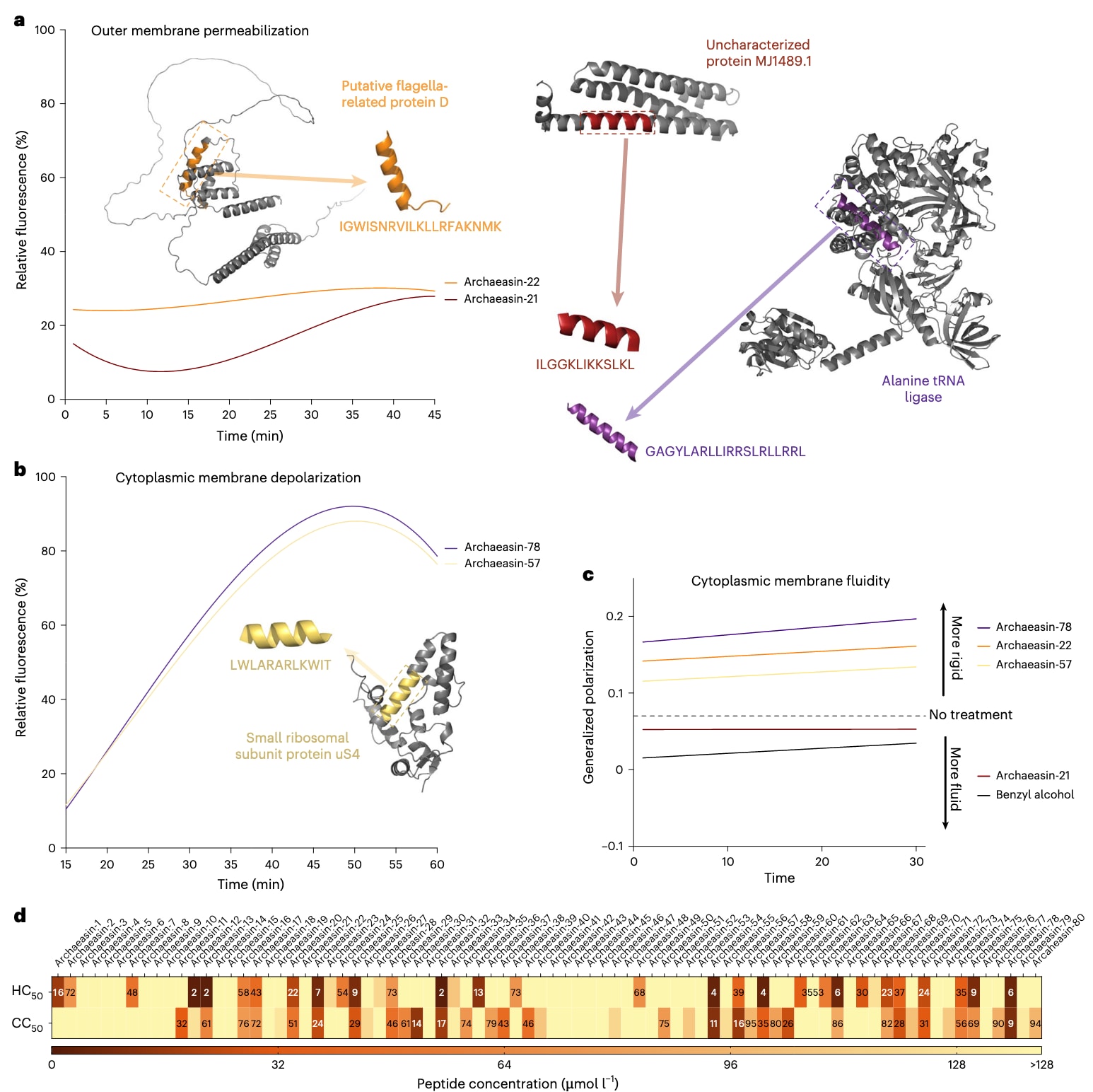
图4 | 古菌来源抗生素的作用机制、溶血活性与细胞毒性 为评估古菌来源的肽类抗生素(EPs)是否作用于细菌膜,对其在 鲍曼不动杆菌 ATCC 19606 上活性的肽进行了外膜通透化与胞质膜去极化实验。a,b 显示了两种表现最佳的古菌素:一种在外膜通透化实验中活性最高,另一种在膜去极化实验中效果最显著(更多结果见扩展数据图6)。实验中使用荧光探针 NPN 来评估外膜通透化(a),使用探针 DiSC3-5 来评估胞质膜去极化(b)。图中数值为两种探针的相对荧光信号,采用非线性拟合,与未处理对照组(缓冲液+细菌+荧光染料)基线比较,并以抗生素 多黏菌素B 与 左氧氟沙星 作为参照。c 显示了经古菌素处理的鲍曼不动杆菌中,Laurdan广义极化值随时间的变化。该指标用于评估古菌素对细胞质膜脂质包装(膜有序性)的影响。结果显示:外膜通透化能力较强的 archaeasin-21 与 archaeasin-22,以及胞质膜去极化作用更显著的 archaeasin-57 与 archaeasin-78,均引起了膜有序性变化。广义极化值升高表示膜刚性增强,下降则表示膜流动性增加。苄醇作为正对照诱导膜流动化,未处理细胞作为负对照。数据为三次独立实验在30分钟内的平均值线性回归结果。d 展示了对红细胞(RBCs)与 HEK293T 细胞的溶血与细胞毒性浓度,即分别导致50%溶血与细胞死亡的浓度(HC50 与 CC50),通过非线性回归拟合剂量-反应曲线获得。所有实验均独立重复三次(扩展数据图7)。a 与 b 面板中的蛋白与肽结构图由 PyMOL 分子图形系统 v3.0 (Schrödinger) 绘制。
2.7 古菌素在临床前动物模型中的抗感染活性
为评估候选古菌素在复杂生理环境中是否仍能保持抗菌效力,研究人员在两种小鼠模型中进行了测试:皮肤脓肿模型和股部深部感染模型(图5a)。在这两个模型中,均使用了鲍曼不动杆菌作为致病菌,该病原体可导致血液、泌尿道、肺部及皮肤创口感染,并因耐药性而成为住院患者死亡的重要原因。
结果表明,三种候选古菌素对鲍曼不动杆菌表现出显著活性且无明显细胞毒性(CC50 < 64 μmol l⁻¹):来自 Aeropyrum pernix 的 archaeasin-2(MIC=4 μmol l⁻¹)、来自 Ignicoccus hospitalis 的 archaeasin-17(MIC=2 μmol l⁻¹)以及来自 Sulfurisphaera tokodaii 的 archaeasin-73(MIC=8 μmol l⁻¹)。
在皮肤脓肿模型中,将
在股部深部感染模型中(图5c),小鼠先接受两轮环磷酰胺处理以诱导免疫抑制,再通过肌肉注射感染
这些体内实验结果支持了古菌素在生理条件下的抗生素特性,为其作为潜在抗菌药物的进一步开发提供了有力依据。
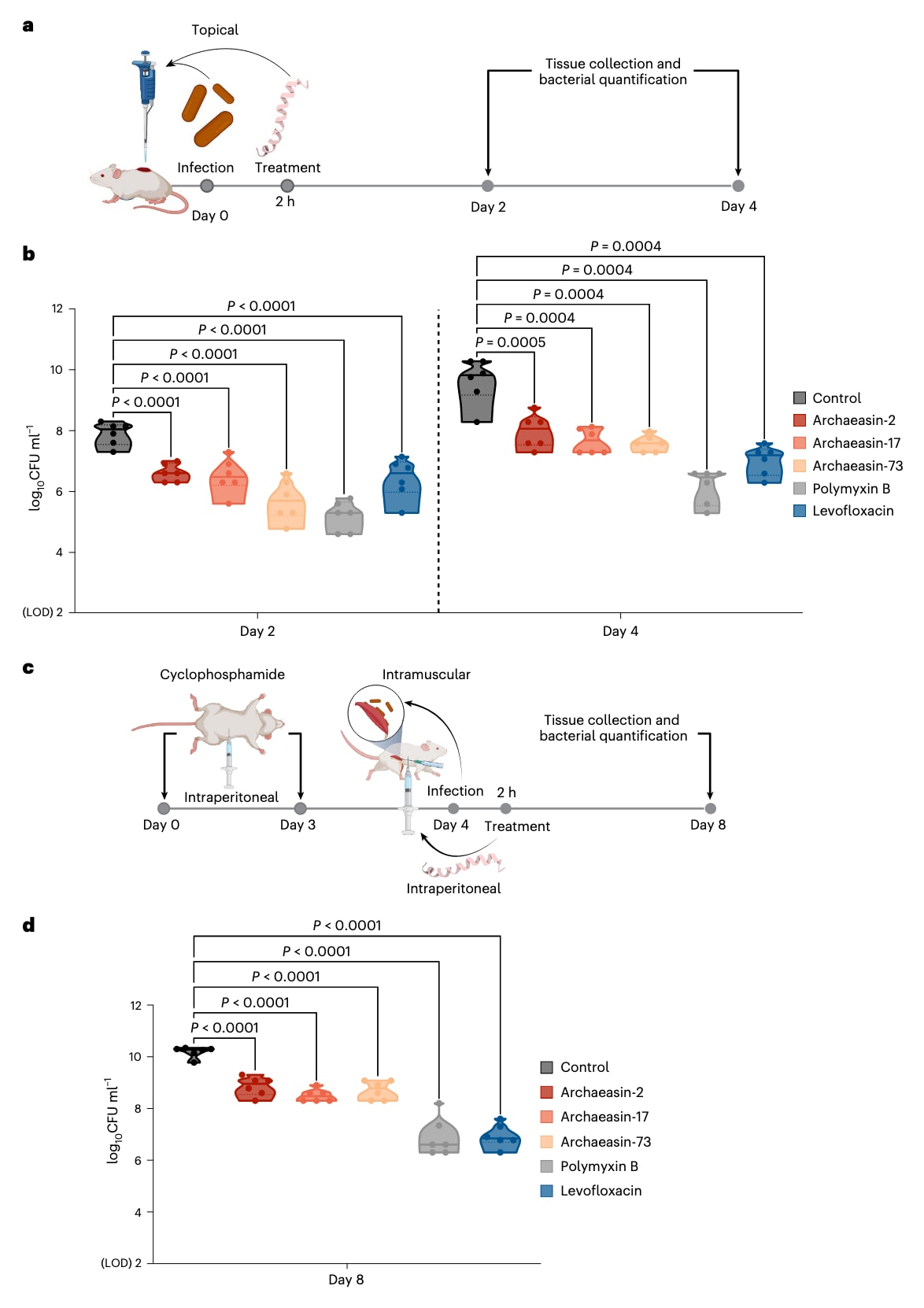
图5 | 古菌素在动物模型中的抗感染活性 a,小鼠皮肤脓肿模型的示意图,用于评估古菌素对 鲍曼不动杆菌 ATCC 19606 的抗感染活性(n = 6)。b,在感染后单次给予 MIC 剂量的 archaeasin-2、archaeasin-17 和 archaeasin-73,均能在处理后4天内抑制感染扩散,与未处理对照组相比显著降低了细菌负荷。尤其是 archaeasin-73,在部分小鼠中将感染水平大幅降低,其活性与对照抗生素多黏菌素B相当。c,中性粒细胞缺乏小鼠的股部感染模型示意图,古菌素通过腹腔注射方式给药。在注射后第4天(即实验第8天),评估其对 鲍曼不动杆菌 ATCC 19606 的抗感染作用(n = 6)。d,结果显示,所有在 MIC 剂量下给予的古菌素均表现出抑菌作用,能够控制感染,但其效力弱于多黏菌素B与左氧氟沙星,与未处理对照组相比差异显著(扩展数据图8)。菌落形成单位(CFU)检测下限为
3 讨论
该研究利用深度学习模型 APEX 1.1 系统性地探索了古菌蛋白质组,揭示了古菌中大量此前未被识别的抗生素分子,凸显了古菌作为抗菌剂新来源的潜力,拓展了以往主要依赖细菌和真菌的天然抗生素研究方向。研究报道了archaeasins这一类具有独特序列多样性的抗菌肽(AMPs),并通过协同实验表明它们能够协同增强抗菌效力。尤其是在 Methanocaldococcus 与 Methanothermobacter 等物种中观察到较低的 FICI 值,提示可以利用这种协同效应开发联合疗法,为治疗多重耐药病原体提供新的方案。
作用机制研究显示,archaeasins 的主要抗菌方式是去极化细菌胞质膜,而非传统 AMPs 常见的外膜通透化。这种去极化会破坏细菌内稳态并导致细胞死亡,其机制与近年来报道的部分小开放阅读框(sORF)编码肽更为接近。
在体内实验中,archaeasins 在小鼠皮肤脓肿与股部深部感染模型中均表现出良好的抗感染活性,尤其是 archaeasin-73,其效力可与传统抗生素多黏菌素B和左氧氟沙星相媲美。这些结果表明,archaeasins 具备成为潜在治疗药物的可能性,特别适用于由耐药菌株如鲍曼不动杆菌引起的感染。更为重要的是,实验未观察到明显毒性,进一步支持了其安全性。
尽管结果令人鼓舞,但仍存在若干挑战与局限。例如,长期疗效与安全性尚需系统评估,包括药代动力学、药效学及潜在免疫原性等。此外,深度学习方法的局限也不可忽视。当前模型基于序列信息,缺乏结构层面的描述,未来引入三维结构特征有望提升预测准确性。为避免模型在 MIC 预测中过度乐观,训练中引入了“非活性”数据并赋予伪MIC值(512 μmol l⁻¹),但这可能带来噪声。未来可通过对比学习等方法减少这一问题。
另一个挑战是古菌蛋白信息的有限性。虚拟筛选仅基于 UniProt 的已审核序列,导致部分类群(如 DPANN 和 Asgardarchaeota)未能被充分表征。未来将拓展至未审核序列以补充分析。此外,该研究仅使用雌性小鼠以保持与既有研究一致,但性别相关的生理差异(免疫反应、激素水平、微生物组等)可能影响感染结果和抗菌效力,这一点在未来研究中应予以关注。
在皮肤感染模型中,古菌素在感染后第2至第4天疗效有所下降,推测可能与其易受蛋白酶降解及体内清除有关。未来可通过化学修饰(如D-氨基酸引入、环化、PEG化)提高稳定性,延长体内疗效。尽管已有研究表明EPs较不易诱导耐药,但仍需进一步检验 archaeasins 的耐药风险。
另外,随机序列对照的研究显示,打乱的抗菌肽序列失去了活性,提示其生物来源的天然序列特征对抗菌效应至关重要。结合以往研究中发现的“隐藏”或“加密”活性肽,该研究进一步支持了一个重要假设:许多此前未被识别的生物活性肽,包括源自人类蛋白质组的肽,可能在宿主免疫与生理过程中发挥关键作用。
综上所述,该研究展示了利用深度学习发掘古菌蛋白质组作为抗生素新资源的前景。archaeasins 的发现为新型抗菌药物的研发奠定了基础,并为应对抗菌耐药性开辟了新的研究方向。