PNAS 2025 | 从头设计高效抑制梭菌家族毒素的分子

获取详情及资源:
0 摘要
艰难梭菌(Clostridioides difficile)仍是医院相关感染的主要病因,其关键毒力因子毒素B(TcdB)会引发严重结肠炎和反复发作的疾病。与之密切相关的Paeniclostridium sordellii来源的毒素TcsL,则会导致一种罕见但常常致命的中毒性休克综合征,尤其发生在妇科和产科环境中。研究报道了通过从头设计获得的小型蛋白质结合物(minibinders),能够直接中和TcdB和TcsL,阻止其进入宿主细胞。
借助深度学习与Rosetta方法,研究者生成了高亲和力结合物,可在皮摩尔级效力下保护细胞免受毒素作用,并且在TcsL的情况下,还能在小鼠致死性毒素挑战实验中显著延长生存期。针对TcdB设计的蛋白表现出在蛋白酶消化及酸性环境中的卓越稳定性,因此极为适合口服递送,这一特性对于局限在胃肠道的艰难梭菌感染尤为重要。而在TcsL研究中,从48个初始设计和48个优化设计中筛选出了高效抑制剂,突显了计算设计在快速开发应对致命细菌毒素策略中的巨大潜力。
1 引言
艰难梭菌毒素B(TcdB)是艰难梭菌感染(CDI)发病的关键驱动因子。这是一种常见且有时致命的结肠医院相关疾病,仅在美国每年就有约50万例病例。与之相关的同一家族毒素TcsL,由Paeniclostridium sordellii产生,可通过全身性中毒性休克引发一种罕见但高度致死的疾病,主要影响产后女性。这些相关毒素导致的不同临床表现,源于它们通过与特异性宿主受体相互作用而表现出的细胞嗜性差异。
目前,急性CDI的标准治疗方案是万古霉素和菲达霉素,但其对肠道菌群的干扰会加剧疾病进程,从而导致较高的复发率,使患者陷入反复发作性CDI(rCDI)的循环。针对TcdB的治疗性单克隆抗体Bezlotoxumab能够使rCDI的复发率下降约10%,为TcdB中和治疗提供了坚实的生物学依据。然而,由于其高昂的成本、复杂的给药途径(静脉注射)以及在急性CDI中的疗效仍不明确,该药仅被推荐用于rCDI高风险人群。同时,目前尚不清楚Bezlotoxumab与口服给药、价格相当且同样降低rCDI复发率的菲达霉素相比孰优孰劣。因此,亟需开发新型抗TcdB治疗手段,要求其可口服、能在感染部位直接发挥作用、成本适宜,并可在暴发期间作为预防性用药,而不会带来明显的耐药风险。
在P. sordellii感染中,其高死亡率反映出针对相关中毒性休克综合征缺乏有效治疗方法。值得注意的是,缺乏TcsL产生的致病菌株在动物模型中几乎不表现出明显的疾病症状,这也验证了TcsL作为预防和治疗毒性休克的关键靶点。因此研究者推测,从头设计的小型蛋白抑制剂或许能为治疗梭菌毒素相关疾病提供新的临床解决方案。与单克隆抗体相比,小型蛋白在生产简便性、热稳定性和抗蛋白酶降解性方面具有显著优势,同时仍能保持对靶点的高亲和力与特异性。研究目标在于设计小型蛋白,精准结合梭菌毒素与细胞表面受体的作用位点,从而阻止毒素进入细胞。
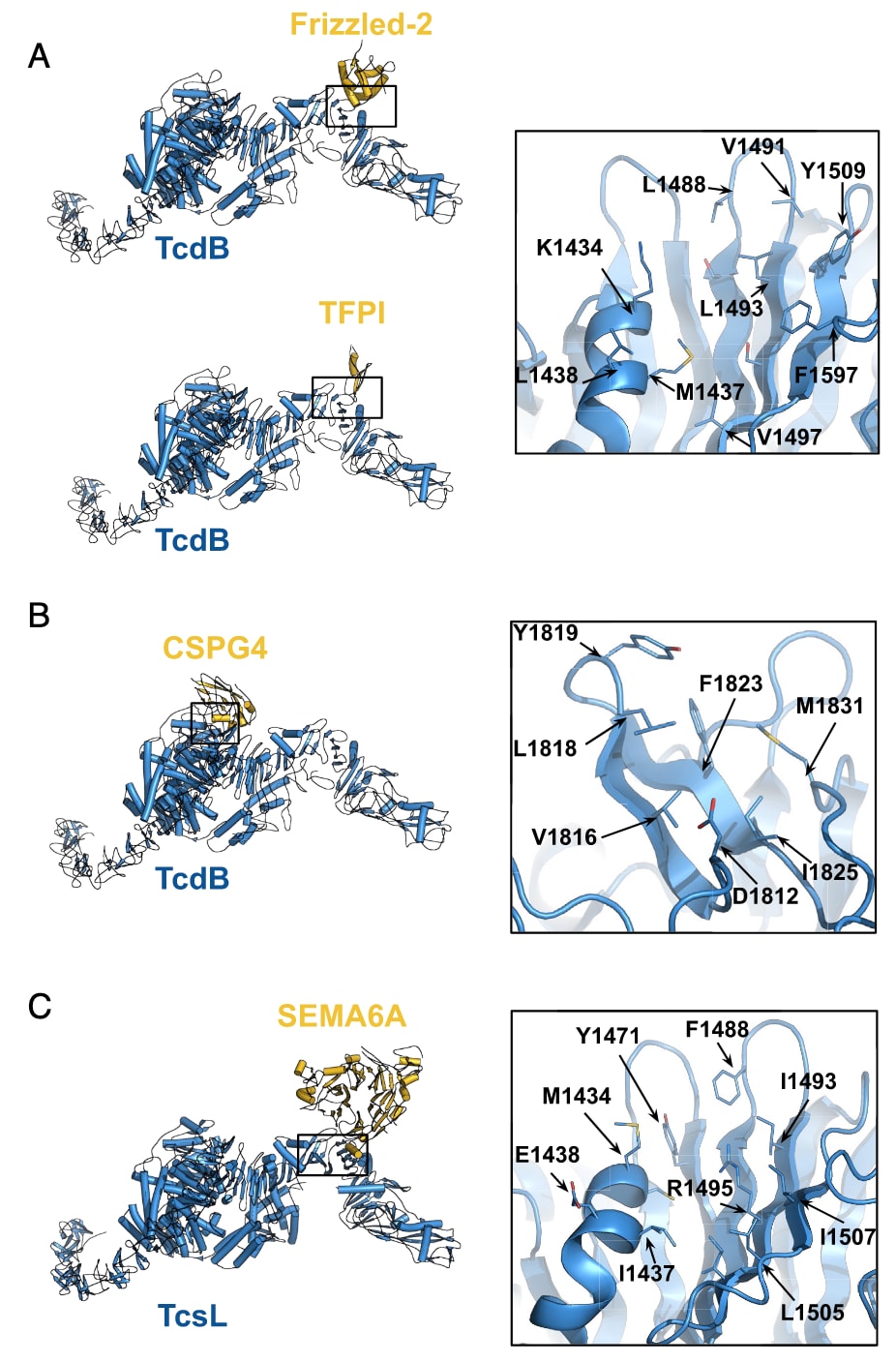
图1 | 梭菌毒素与宿主细胞受体复合物的复合模型 内嵌框标注了在设计过程中所针对的残基。(A) TcdB [PDB 6OQ5] 与 Frizzled-2 [PDB 6C0B] 及 TFPI [PDB 7V1N] 的复合物。(B) TcdB [PDB 6OQ5] 与 CSPG4 [PDB 7ML7] 的复合物。(C) TcsL [PDB 8JB5] 与 SEMA6A [PDB 6WTS] 的复合物。
2 Frizzled/TFPI结合位点的抑制
TcdB在不同变体中表现出不同的受体使用模式。该毒素在远端RBD上具有一个受体结合位点,可与Frizzled-1/2/7或TFPI相互作用;另一个结合位点位于毒素核心区,能够结合宿主CSPG4,并且在TcdB2的情况下可独立于Frizzled发挥作用。研究首先针对TcdB的Frizzled/TFPI结合界面,建立了整合Rosetta与深度学习方法的设计流程。通过RifGen和RifDock从三螺旋与四螺旋束库中生成构象,重点聚焦结合界面的疏水残基,随后使用ProteinMPNN分配序列,并结合AlphaFold2进行初步筛选。最终共获得约15,000个设计,并利用酵母表面展示进行结合筛查,随后通过位点饱和突变进行优化。
经过序列优化后,亲和力最高的结合体在大肠杆菌中表达,并形成三组单分散SEC峰的设计,分别命名为group 1、group 2和group 3。其中group 2的结合体表现出极高的稳定性,即使解离时间超过1小时仍几乎未检测到解离,提示其
然而,TcdB在结肠腔中发挥作用,因此抑制剂若要用于治疗,必须能够在胃肠道中抵抗广泛的蛋白酶降解。实验表明这些设计在模拟肠液中降解迅速。为提高蛋白酶稳定性,研究人员在group 1和group 2的设计中引入了二硫键,其中group 2的二硫键连接第一、二螺旋之间的环与C端,有效地“锁定”了整体构象。结果显示,二硫键稳定化的设计不仅保持了结合活性,而且在37℃下的模拟肠液中可稳定1小时不降解。尤其是**ssfzd84(group 2, C18-57二硫键修饰)**几乎未发生降解,仅在C端标签部分丢失,同时在中和实验中表现出更强效力,IC50仅为24 pM。
这一系列结果表明,针对Frizzled/TFPI结合界面的理性设计能够产生高亲和力、结构稳定且具有潜在临床应用价值的TcdB抑制剂。
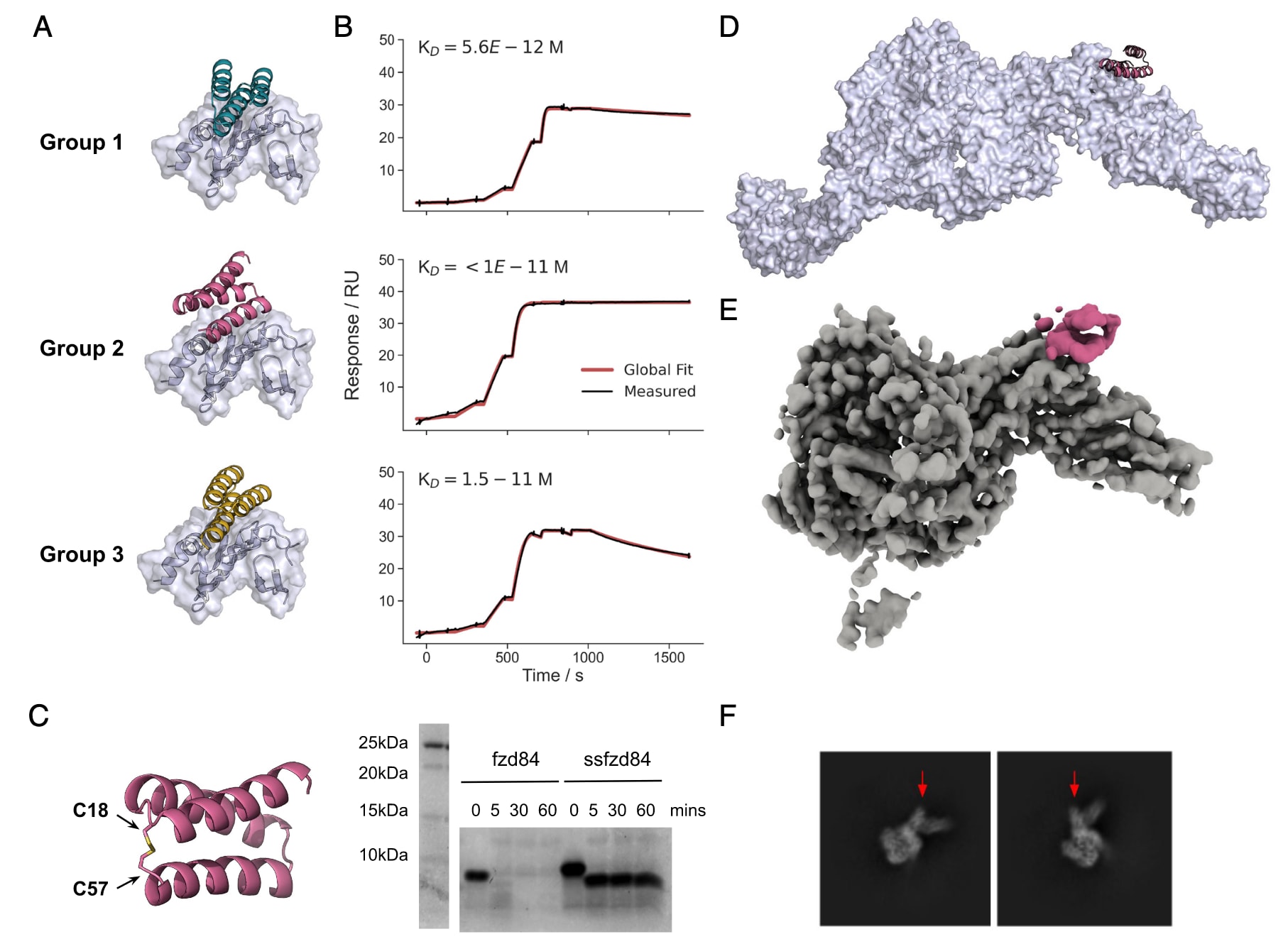
图2 | 抗TcdB Frizzled阻断型小型结合体的设计 (A) 三类高亲和力小型结合体家族的设计模型。(B) 三个代表性设计(fzd13,来自group 1;fzd48,来自group 2;fzd24,来自group 3)的单循环动力学分析。实验条件为:TcdB RBD固定在芯片上,起始浓度100 nM,6步4倍梯度稀释。(C) fzd84小型结合体的二硫键稳定化设计。左图为二硫键引入位置;右图为Coomassie染色的SDS-PAGE结果,显示其在模拟肠液(含0.1 mg/mL胰蛋白酶与糜蛋白酶)中37℃处理后的时间依赖性降解情况。(D) fzd48与全长TcdB(PDB: 6OQ5)复合物的设计模型。(E) fzd48(粉色)与TcdB复合物的冷冻电镜分割密度图。(F) 冷冻电镜的典型类别平均图像,箭头标注了因fzd48结合而产生的额外密度信号。
3 CSPG4结合位点的抑制
研究人员将同样的设计策略应用于CSPG4结合位点,但由于该位点拓扑结构复杂,难度更高。通过RifGen/RifDock结合ProteinMPNN与序列优化,并在设计中引入单一二硫键,最终获得了两类设计(group 1与group 2),其结合力达到低纳摩尔或亚纳摩尔水平。这些设计在使用依赖CSPG4进入细胞的TcdB2时,能够有效中和毒素,在0.1 pM TcdB存在的条件下,cspg18、cspg27与cspg35的IC50分别为238 pM、520 pM与332 pM,表现出显著的保护作用。
尽管这些分子已含有一个二硫键,但仍容易被蛋白酶降解。通过引入第二个二硫键,设计获得了显著的稳定性提升:在模拟肠液中处理1小时后仍保持完整,而其单二硫键版本在5分钟内即被完全降解。重要的是,这种增强稳定性并未削弱其中和效力。进一步的冷冻电镜结构解析显示,group 2结合体cspg67与TcdB复合时,第二与第三螺旋与邻近CSPG4结合槽的疏水斑块(V1816、L1818、Y1819、F1823与M1831)相互作用,而连接这两条螺旋的环则延伸进入结合槽,第二螺旋C端通过K32与R36与TcdB的D1812形成极性作用。由于该结合位点位于GTD与CPD核心区域而非远端DRBD,解析结果得以清晰辨认大侧链残基(F42与W45),并与设计模型高度一致。
为进一步探究Frizzled阻断型与CSPG4阻断型小结合体联合使用的效果,研究者在Vero细胞上进行了梯度联合实验。在依赖双受体(TcdB1, EC99=0.1 pM)的系统中,ss2cspg18与ssfzd84联合应用表现出协同效应:随着ssfzd84浓度增加,曲线明显左移,IC50由520 pM下降至35 pM,同时保护率提升至100%,远高于单独使用ss2cspg18的情况。而在单CSPG4依赖系统(TcdB2, EC99=0.1 pM)中,增加ssfzd84并未带来额外优势。
这些结果表明,针对CSPG4结合位点的理性设计可获得高亲和力且稳定的抑制剂,而其与Frizzled阻断分子联合使用时,能够实现协同中和效应,为多受体依赖型毒素的治疗提供了更强的保护潜力。
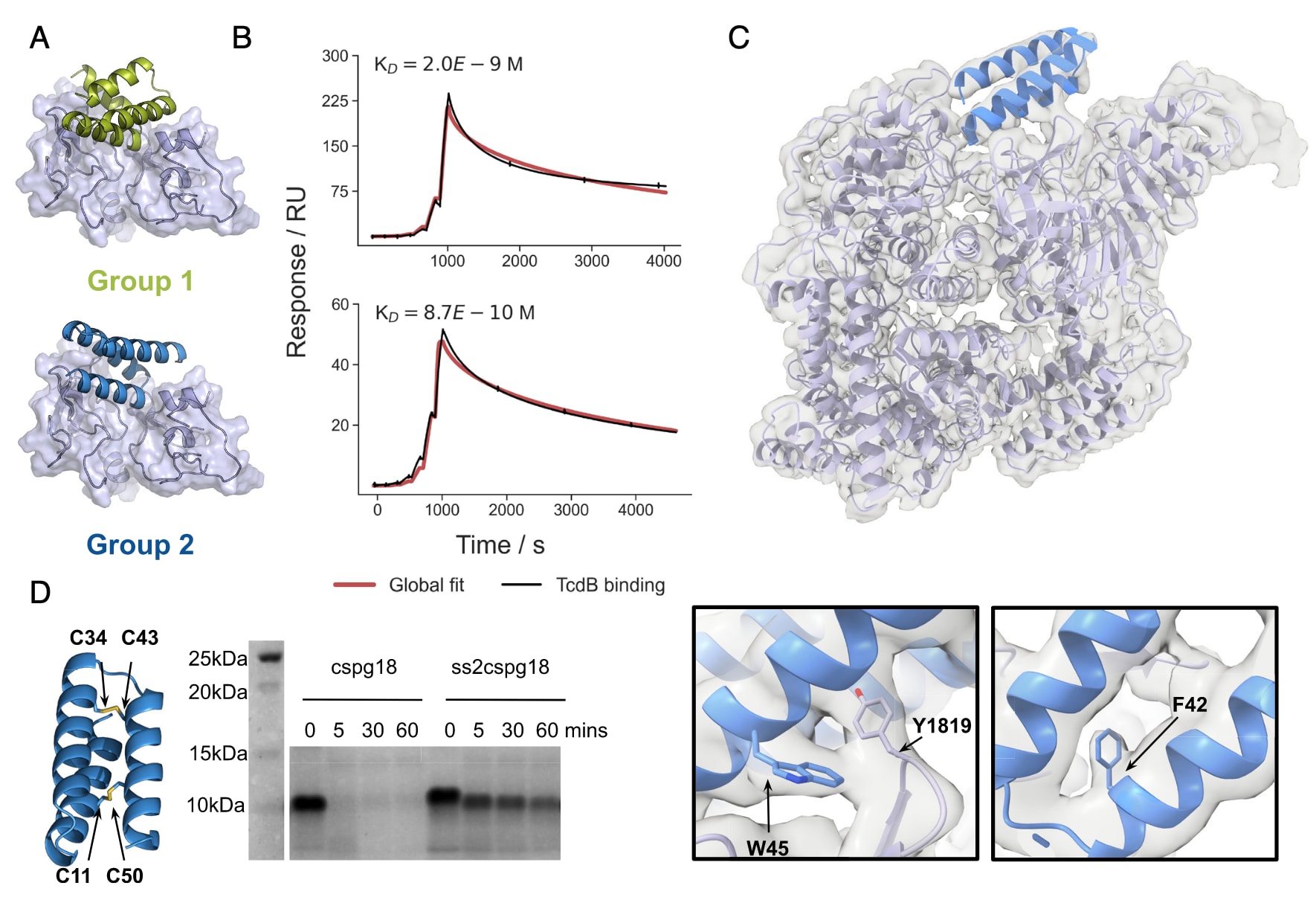
图3 | 抗TcdB CSPG4阻断型小型结合体的设计 (A) 两类CSPG4阻断型小结合体家族的设计模型。(B) group 1 (cspg4)与group 2 (ss2cspg18)设计的单循环动力学分析。实验中将小结合体氨基偶联到芯片表面,与全长TcdB进行6步4倍梯度稀释结合实验,最高浓度100 nM。(C) 设计模型对接至冷冻电镜密度图,结果显示观测密度与设计模型高度一致。下方插图突出显示了在靶标–结合体界面解析出的关键侧链残基。(D) CSPG4阻断型小结合体的二硫键稳定化设计。左图显示了在ss2cspg18中引入的两个二硫键位置;右图为Coomassie染色的SDS-PAGE结果,比较cspg18与双二硫键版本(ss2cspg18)在模拟肠液(含0.1 mg/mL胰蛋白酶与糜蛋白酶,37℃)中的时间依赖性降解情况。
4 fzd/cspg融合构建体在益生酵母S. boulardii中的分泌
在作用部位原位生产生物制剂具有显著优势,因为它能实现持续的治疗分子供应,而不必依赖频繁的口服给药。益生酵母Saccharomyces boulardii是一种能够在下消化道短暂定植的菌株,与其他实验性细菌递送平台不同,它对抗生素不敏感,并已被成功用于向肠道递送抗体样分子。基于这一特性,研究团队尝试利用S. boulardii生产并分泌TcdB抑制性小结合体(minibinders)。
在体外实验中,二硫键稳定化的单体小结合体能够以高达50 nM的浓度被分泌。更为重要的是,同时阻断CSPG4和Frizzled的融合小结合体(ss2cspg18与ssfzd84通过(G4S)₄连接)也能够被S. boulardii高效分泌。培养上清液对TcdB1表现出强效中和作用,IC50仅为22 pM。相比之下,当两者仅通过过短的GSG连接(不足以同时结合TcdB上的两个受体界面)时,中和效力明显下降,IC50升高至672 pM。这表明,通过连接不同结合域并实现多价效应(avidity),能够显著增强抑制活性。
这些结果证明,多种格式的小结合体都可以通过S. boulardii实现高效生产与分泌,为利用小型蛋白构建的合成生物制剂在多种胃肠疾病中的应用奠定了基础。
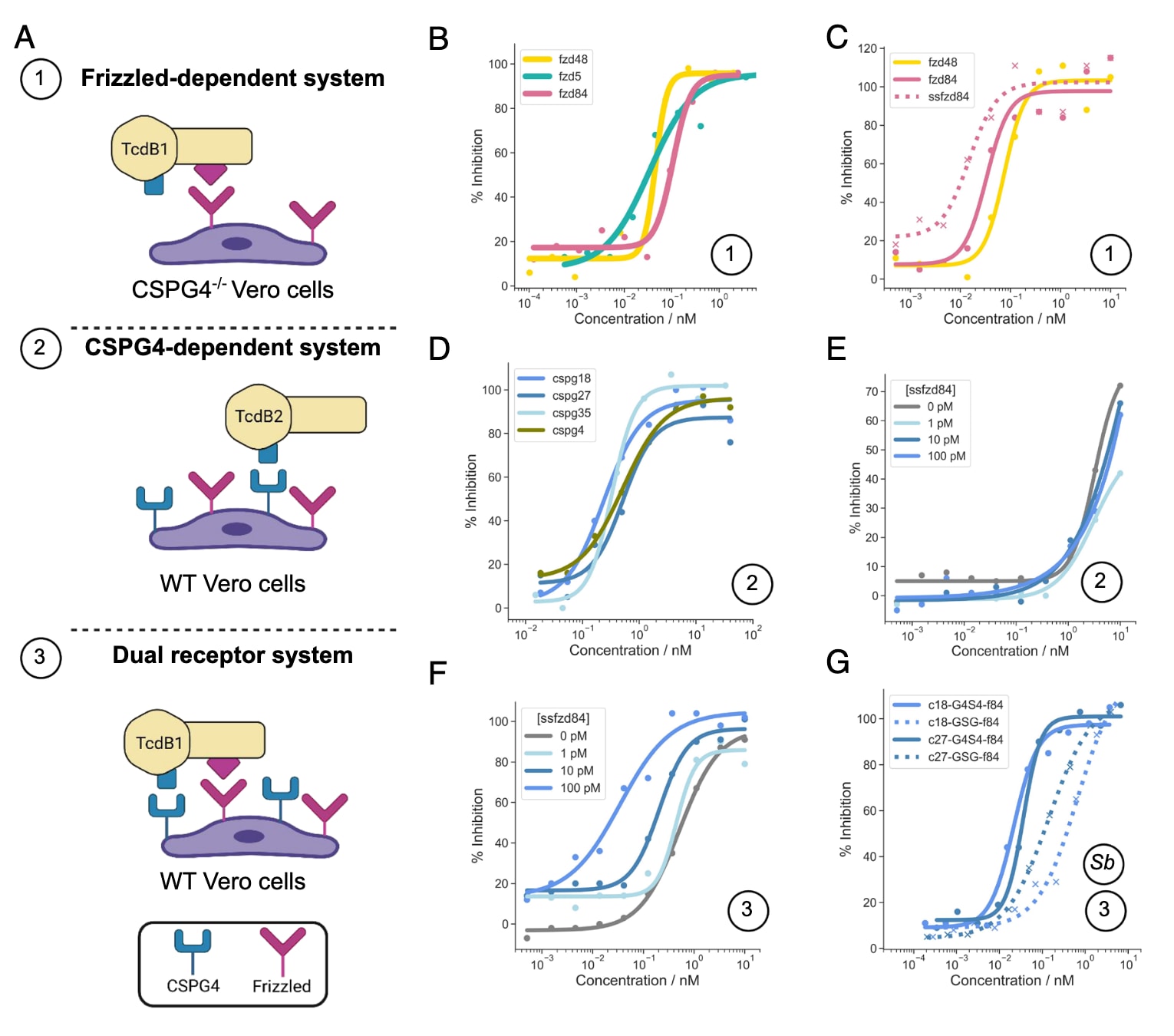
图4 | 小结合体在Vero细胞中对TcdB的中和作用 (A) 三种实验体系的示意图,用于评估不同类别小结合体单独或联合的中和活性。(B) 优化后的Frizzled结合体,包括group 1的fzd5和group 2的fzd48、fzd84。(C) 蛋白酶耐受型的序列优化设计ssfzd84,与蛋白酶敏感型设计的比较。(D) 优化后的CSPG4结合体,包括group 1的cspg4和group 2的cspg18、cspg27、cspg35。(E) 在CSPG4依赖系统中,固定不同浓度的ssfzd84(0、1、10、100 pM),并对ss2cspg18进行梯度滴定。(F) 在双受体依赖系统(Frizzled+CSPG4)中,同样以不同浓度的ssfzd84配合ss2cspg18进行梯度滴定。(G) 由S. boulardii分泌的Frizzled-CSPG4融合结合体的中和活性。展示了两种融合构建体:一种为ss2cspg18 (c18)或ss2cspg27 (c27)与ssfzd84 (f84)通过短GSG连接肽融合(预期无法同时结合两受体位点,虚线表示),另一种为通过长(G4S)₄连接肽融合(可同时结合两个靶点,实线表示)。所有结果均为独立重复实验中的代表性示例,IC50数值为多次实验的平均值。相同坐标轴下绘制的曲线均来自同一实验。图中圆圈编号对应(A)中的实验体系,Sb表示使用S. boulardii分泌的蛋白。
5 TcsL的中和与致死性毒性休克的防护
在成功中和TcdB的基础上,研究进一步尝试针对P. sordellii致死毒素TcsL设计抑制剂。TcsL可引发人类及家畜的高度致死性疾病,在人类中主要见于产后或流产相关病例,后者常与米索前列醇/米非司酮的使用有关。与TcdB不同,TcsL通过单一已知的受体结合位点与宿主细胞表面的SEMA6A/SEMA6B结合,其结构方式与TcdB-Frizzled相似。
研究利用生成式AI方法RFdiffusion,从随机氨基酸坐标出发,经过50步去噪生成55–65残基的小型蛋白结合体,并针对SEMA6A界面的疏水热点进行设计。经过ProteinMPNN生成序列并结合AlphaFold2筛选后,共获得10,000条轨迹并缩减为48个候选在大肠杆菌中表达。最高亲和力设计F4的
在HCT116细胞中(50 pM TcsL, EC99),B4与B10的IC50分别为238 pM与210 pM。在SEMA6A高表达的HULEC5a细胞中(0.5 pM TcsL, EC90),两者同样表现出高效中和(IC50分别为248 pM与157 pM)。尽管TcsL与TcdB序列同源性达76%,这些设计对TcdB并无结合活性,表现出高度特异性。
由于小结合体在体内循环时间短,研究进一步将B4与白蛋白结合域M79融合以延长半衰期。该融合体不影响结合活性或中和效力。在小鼠实验中,B4-M79(0.06 mg/kg,毒素的1000倍摩尔过量)与TcsL共同给药,显著延长存活时间,从对照组中位数4 h延长至8 h。重复给药实验中,在预防性方案下,接受B4-M79的小鼠全部存活且几乎无肺水肿;在更严格的治疗性方案下,10只小鼠中有8只存活至实验结束(14 h),其余2只于11.5 h出现症状死亡,而对照组中位生存时间仅为5.5 h。
此外,B4-M79可冻干保存而不损失中和活性,为其长期储存和临床转化提供了可行路径。
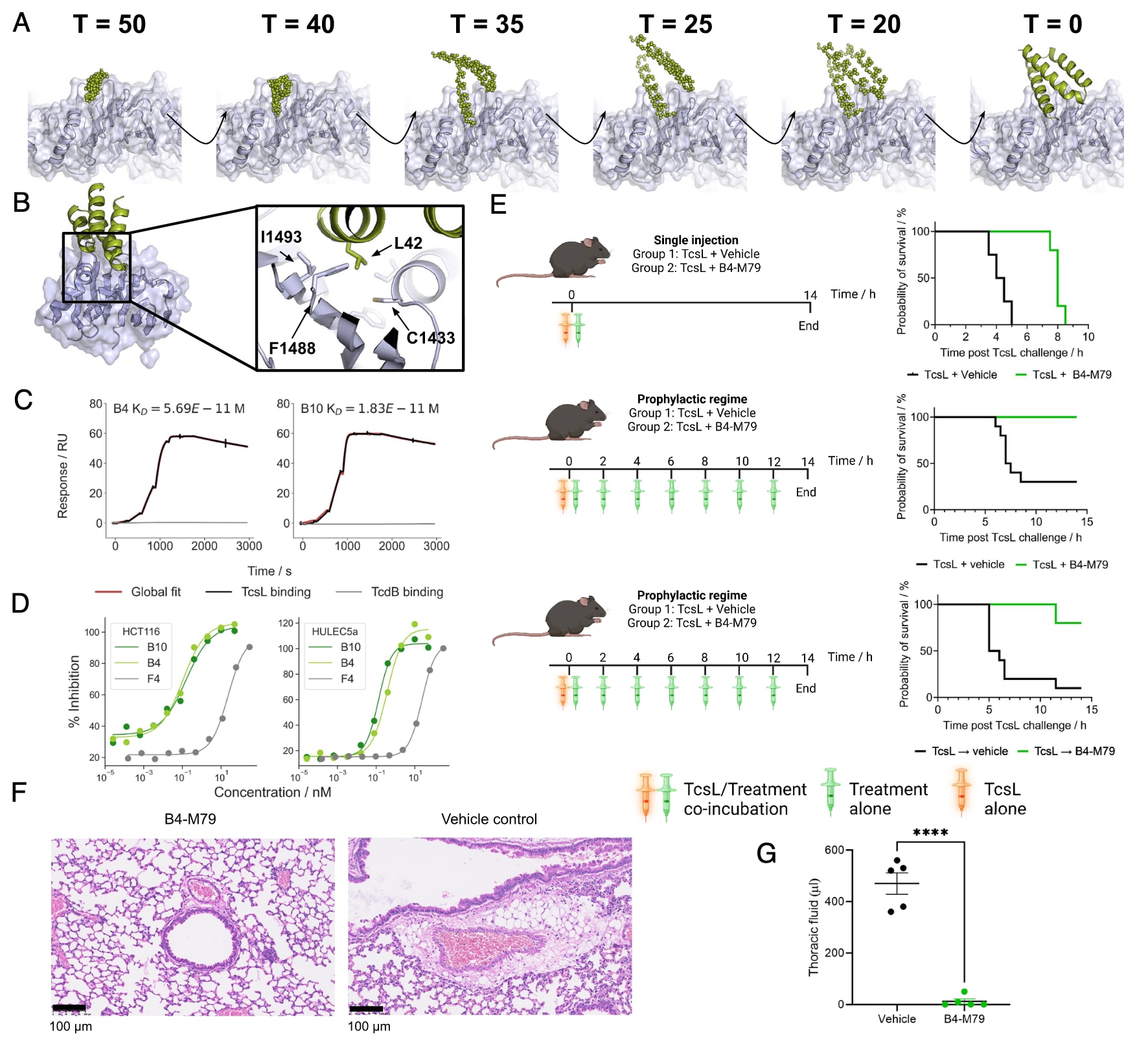
图5 | TcsL阻断型小结合体的设计与功能验证 (A) RFdiffusion去噪轨迹:从随机噪声起始(T=50)逐步去噪至完全成型的设计(T=0)。(B) F4结合体与TcsL RBD的设计模型,插图框突出显示了界面主要由疏水口袋构成。(C) 单循环动力学分析:利用SPR检测优化设计(B4与B10)对TcsL(黑色)和TcdB(灰色)的结合活性。实验条件为毒素RBD固定,起始浓度100 nM,6步4倍稀释系列。(D) TcsL中和实验:亲本设计F4及优化设计B4、B10在HCT116细胞(左)与HULEC5a细胞(右)中的中和效力。IC50数值见正文,均为独立重复实验的平均值,曲线为同一实验的代表性结果。(E) 小鼠生存实验:
-
单次给药:TcsL单独或与B4-M79(1000倍摩尔过量)共给药,存活时间由中位4.25 h延长至8 h,P=0.0027(上图)。
-
预防性方案:TcsL与B4-M79共给药并每2小时重复给药,P=0.0002(中图)。
-
治疗性方案:先给TcsL,1小时后再给B4-M79并每2小时重复给药,P=0.0012(下图)。
(F) 小鼠肺部组织学切片:H&E染色显示对照组小鼠肺水肿明显,而B4-M79处理组无显著病理特征。
(G) 肺内积液定量:对照组与B4-M79治疗组的小鼠肺积液比较,线条表示平均值±标准误差;双尾非配对t检验,P<0.0001。
6 讨论
该研究设计的小结合体为应对梭状芽孢杆菌毒素所带来的重大健康挑战提供了一条新途径。与直接杀灭病原体不同,中和毒素的治疗策略可减弱抗菌耐药性的进化压力,因为毒素通常并非病原体存活所必需。针对C. difficile特有的毒力因子尤具吸引力,因为抗生素引起的微生物群紊乱往往加剧疾病进展并诱发复发。通过TcdB中和实现的有效治疗不仅有望改善临床结局,同时也有助于抗生素管理。贝洛妥单抗作为TcdB单克隆抗体已显示出减少复发的疗效,但其临床应用因高成本而受限,并已被停产。
所设计的小结合体靶向Frizzled与CSPG4两个不同结合位点,具有高亲和力、耐蛋白酶降解,并可通过益生酵母S. boulardii分泌实现递送。在融合后,这些分子在双受体依赖的体外实验中表现出pM级IC50的强效中和。小结合体既可作为重组蛋白制备,也可由S. boulardii在肠道原位分泌,具备口服递送和低成本治疗的潜力。未来关键在于其是否能在胃肠环境中保持活性。该研究在高浓度胰蛋白酶和糜蛋白酶条件下模拟实验所得结果显示,其稳定性具有应用前景;后续将进一步在体内验证,联合现有一线抗生素(万古霉素、菲达沙星)进行评估。尽管目前尚无证据表明小结合体在人体会引发强烈免疫反应,但仍需针对急性感染背景下的免疫原性进行检测。
与纳米抗体、DARPins或双特异性抗体类分子相比,该研究方法的优势在于:结合位点可通过计算设计预设,避免了筛选过程中克隆对非关键表位的偏好;同时,结构上以明确的二级结构单元介导结合,而非依赖柔性环,从而更易赋予蛋白酶稳定性。近年来,设计型小结合体已逐渐能够靶向复杂表面拓扑结构,克服了部分天然框架分子结构局限。
该研究进一步展示了深度学习驱动的蛋白设计如何在短时间内(96个候选即可)筛选出高亲和力抑制剂,并具备体内疗效。针对TcsL的设计不仅展现了在小鼠中防护毒性休克的能力,还能耐受冻干处理,为长期储存与应用提供了可能。在P. sordellii感染中,现有治疗主要依赖病灶切除与广谱抗生素,但病死率仍高达约70%,凸显了有效治疗手段的缺乏。该研究小结合体的体内防护效果与良好稳定性,表明其在治疗这一罕见且致死性疾病方面具有显著潜力。
综上,这一策略验证了小结合体在中和梭菌家族毒素中的可行性。在CDI的背景下,需要确保小结合体在作用部位保持足够活性,并且S. boulardii平台能够在体内分泌足量分子。展望未来,这种结合体设计方法不仅可扩展至其他感染相关毒素,也可应用于病原体生物膜、炎症因子等靶点,显示出广阔的应用前景。