Science 2025 | 裸鼹鼠中由cGAS介导的机制可增强DNA修复并延缓衰老
今天介绍的是 Science 期刊最新研究: 裸鼹鼠中由 cGAS 介导的机制可增强 DNA 修复并延缓衰老。裸鼹鼠(Heterocephalus glaber)以高达约37年的寿命成为地球上最长寿的啮齿动物,其独特的抗衰老与抗癌能力长期吸引生物学界关注。最新发表在 Science 的研究揭示,裸鼹鼠在进化过程中通过改变 cGAS(环鸟苷酸–腺苷酸合成酶)功能,实现了对 DNA 修复机制的强化。与人类和小鼠中会抑制同源重组修复(HR)的 cGAS 不同,裸鼹鼠的 cGAS 反而增强了 DNA 修复效率。这一功能逆转仅由 四个关键氨基酸残基(E、S、T、Y) 的变化所造成,使其在 DNA 损伤后维持低水平的泛素化,从而更长时间地结合染色质。这种持久结合促进了 FANCI–RAD50 复合体的形成并加速修复反应。进一步实验表明,携带裸鼹鼠 cGAS 的果蝇寿命显著延长,老年小鼠则表现出衰老标志减轻、毛发变白减少与系统炎症下降。这些效应在突变上述四个氨基酸后完全消失。该研究不仅揭示了长寿物种通过优化 DNA 修复负调控因子实现延寿的分子逻辑,也为未来抗衰老干预与健康寿命延长提供了新的生物学靶点。
Chen, Y.; Chen, Z.; Wang, H.; Cui, Z.; Li, K.-L.; Song, Z.; Chen, L.; Sun, X.; Xu, X.; Zhang, Y.; Tan, L.; Yuan, J.; Tan, R.; Luo, M.-H.; Sun, F.-L.; Liu, H.; Jiang, Y.; Mao, Z. A cGAS-Mediated Mechanism in Naked Mole-Rats Potentiates DNA Repair and Delays Aging. Science 390 (6769), eadp5056. https://doi.org/10.1126/science.adp5056.
0 摘要
高效的DNA修复能力或许是裸鼹鼠长寿的重要原因。然而,目前尚不清楚这种动物是否具备针对DNA修复抑制因子进行功能优化的独特机制。研究发现,裸鼹鼠的环鸟苷酸–腺苷酸合成酶(cGAS)在进化过程中通过四个氨基酸残基的改变,失去了人类与小鼠同源蛋白在同源重组修复(HR repair)中的抑制作用。这些氨基酸变化削弱了cGAS与E3泛素连接酶TRIM41及分离酶P97的相互作用,从而使其在DNA受损后能够更长时间地滞留于染色质上。这种延长的结合促进了修复因子FANCI与RAD50之间的相互作用,并进一步增强RAD50在损伤位点的募集,最终提升了同源重组修复效率。此外,这四个关键氨基酸还赋予cGAS抗细胞与组织衰老的能力,从而延缓机体老化并延长寿命。因此,调控cGAS的功能可能成为延长寿命的一种潜在机制。
1 引言
裸鼹鼠(Heterocephalus glaber)是一种极端长寿的啮齿动物,寿命可达约37年,比体型相近的啮齿类动物长约一个数量级。其转录组与蛋白编码序列与人类相比更为接近,而非小鼠,这使其成为研究“成功衰老”分子机制的理想模型。基因组稳定性在不同物种的长寿进化中起着关键作用,但目前对裸鼹鼠在DNA修复层面的分子适应性了解仍十分有限。
DNA双链断裂(DSBs)是最具破坏性的DNA损伤类型,其积累与衰老密切相关。同源重组(HR)修复是修复DSB的主要通路之一,其缺陷会导致小鼠与人类出现早衰现象。与HR密切相关的还有Fanconi贫血(FA)修复通路,该途径负责修复DNA链间交联;其中调控因子FANCI除参与FA修复外,还独立参与复制压力响应。因此,FA因子在多种DNA修复类型中可能具有作用。
环鸟苷酸–腺苷酸合成酶(cGAS)是胞质DNA的主要传感器,能识别外源或内源DNA并催化生成环状GMP-AMP(cGAMP),从而激活STING介导的先天免疫反应。然而,在人类与小鼠中,cGAS也定位于细胞核,能抑制HR修复,从而可能促进肿瘤形成并缩短寿命。虽然在长寿进化中,多个DNA修复正调控因子(如SIRT6与BRCA1)被正向选择,但尚不清楚裸鼹鼠的cGAS是否也具有类似抑制作用。
2 裸鼹鼠cGAS增强HR修复以维持基因组稳定
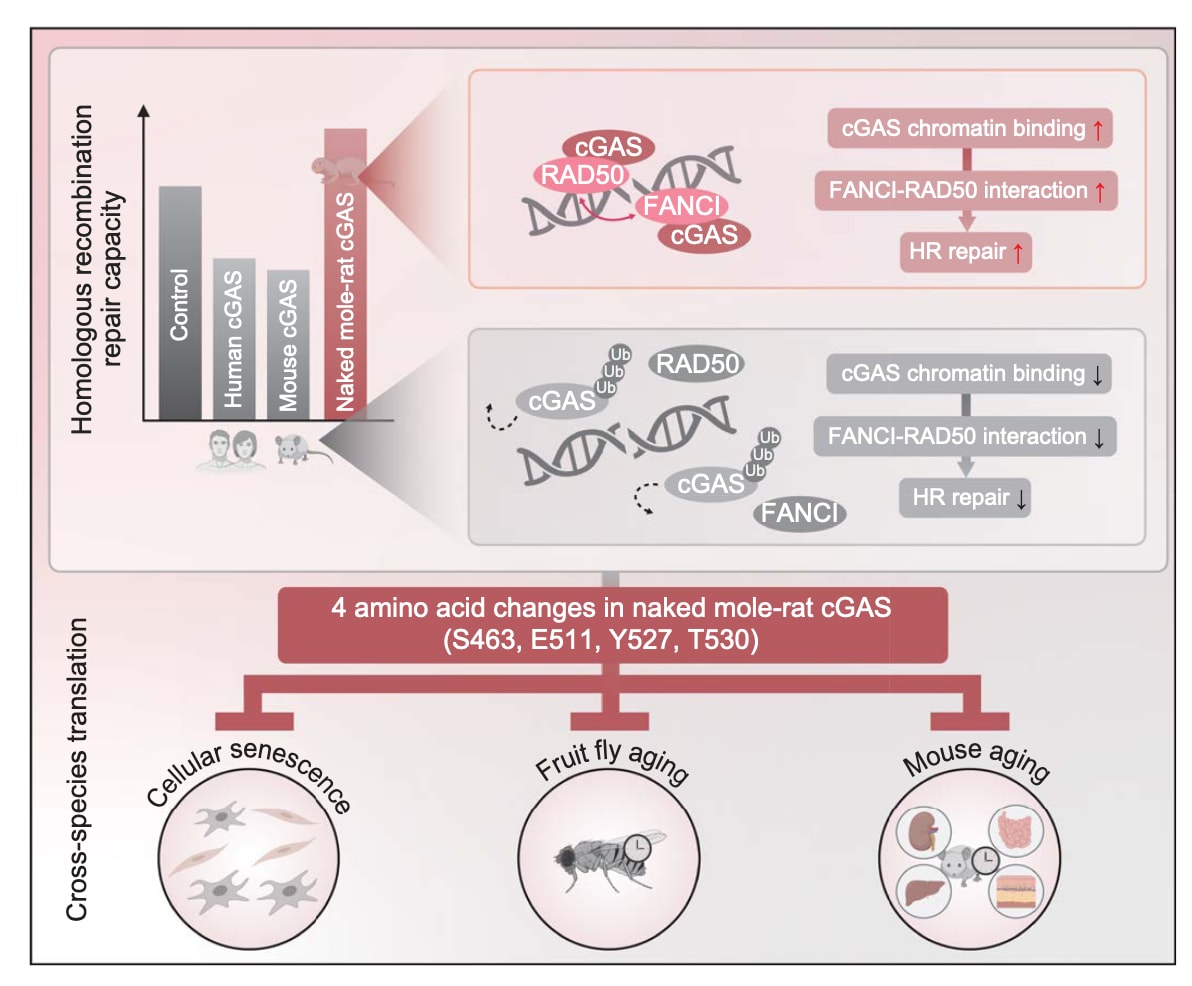
研究者从裸鼹鼠成纤维细胞中克隆出cGAS基因。实验证实其仍具典型的免疫调控功能,但催化活性较人类与小鼠cGAS降低。裸鼹鼠cGAS可被招募至微束辐照诱导的DNA损伤位点,无论在裸鼹鼠还是人类细胞中均可观察到,提示其可能参与DNA损伤响应。
利用GFP报告体系评估HR修复效率发现:人类与小鼠cGAS的过表达会抑制HR修复,而裸鼹鼠cGAS则呈剂量依赖性增强HR修复活性。此外,在裸鼹鼠细胞中通过CRISPR-Cas9敲除cGAS可抑制HR修复,而再表达则恢复修复能力,进一步证明其功能与人、小鼠相反。值得注意的是,即使催化失活突变体(E257A/D259A或D351A)也能促进HR修复,说明这种作用与其酶活性无关。由于HR仅在细胞周期S与G2期发生,研究确认这一促进作用并非源于细胞周期分布变化。免疫荧光结果显示,裸鼹鼠cGAS过表达可加速HR关键重组酶RAD51的募集,并促进DSB标志物γH2AX的清除。彗星实验进一步证实其能增强基因组稳定性,而cGAS缺失则导致DNA稳定性下降。
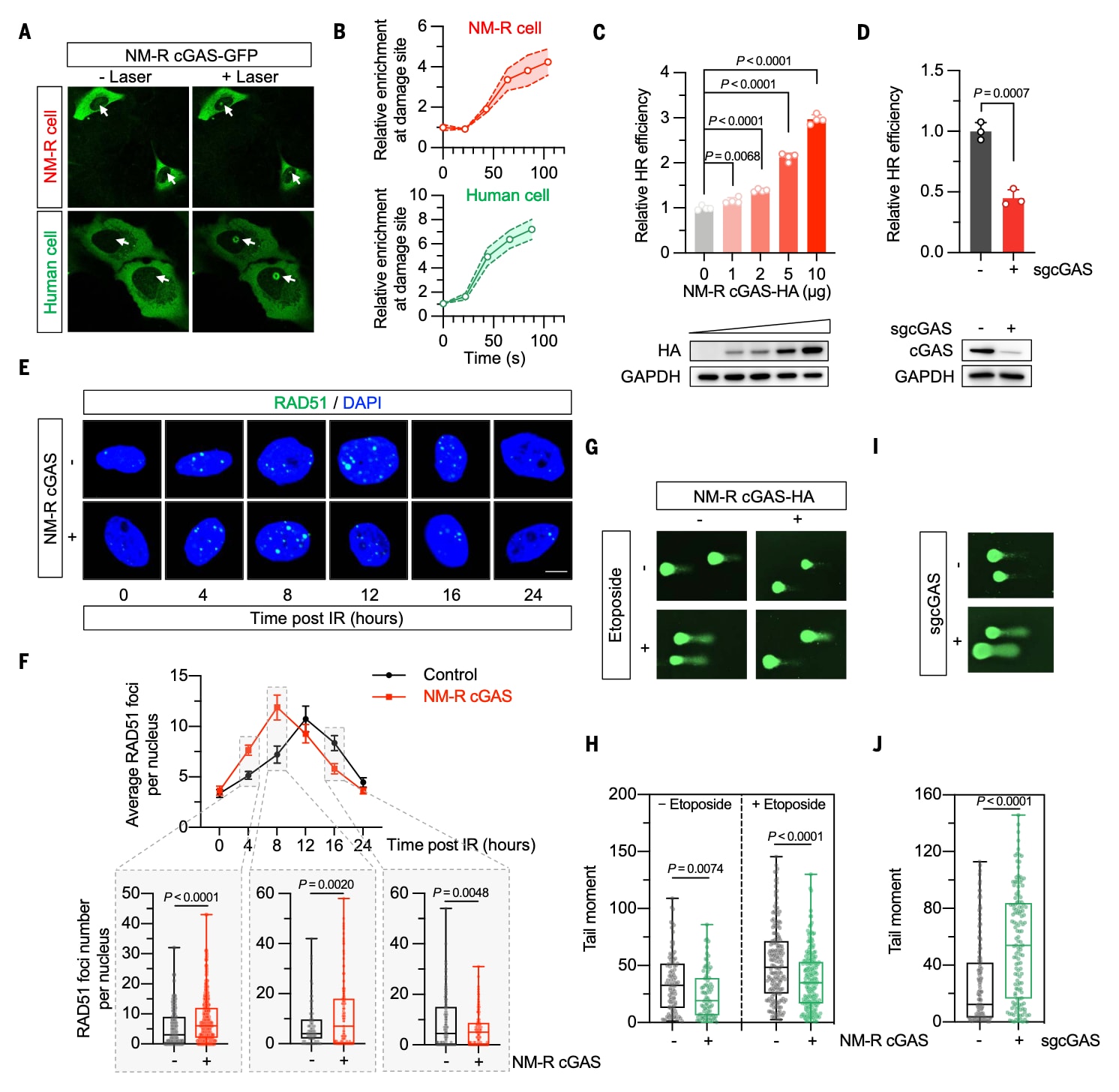
图1|裸鼹鼠cGAS通过促进同源重组修复(HR)维持基因组稳定性 (A、B) 裸鼹鼠cGAS在DNA损伤位点的招募。在人源U-2 OS细胞与裸鼹鼠成纤维细胞(NSFs)中,均观察到带有GFP标签的裸鼹鼠cGAS(图中标注为“NM-R”)被招募至DNA损伤区域。(A)为代表性显微图像,箭头指示细胞受照射的位置;(B)为荧光强度定量分析,阴影区域表示标准误差(SEM)。(C) 在NSFs中裸鼹鼠cGAS增强HR修复效率。细胞共转染含有I-SceI线性化HR报告系统的质粒与不同量的NM-R cGAS-HA表达载体,同时转染DsRed质粒以作对照。cGAS-HA的表达通过Western blot验证。(D) cGAS敲除后HR修复效率下降。在共转染Cas9与cGAS靶向单导RNA(sgRNA)的NSFs细胞群中,HR修复效率显著降低;cGAS的缺失同样经Western blot确认。(C)与(D)中误差线表示标准差(SD),两组间差异通过Student’s t检验评估显著性。(E、F) RAD51募集分析。在是否过表达NM-R cGAS的NSFs中,使用8 Gy X射线照射后,于不同时间点进行免疫荧光染色(比例尺:5 μm)。结果显示,NM-R cGAS过表达显著加快RAD51在损伤位点的聚集;误差线表示标准误差(SEM)。(G、H) 彗星实验检测基因组稳定性(cGAS过表达)。在过表达NM-R cGAS的NSFs中,分别处理或不处理50 μM依托泊苷(etoposide)2小时后收集细胞。结果显示,cGAS过表达显著降低DNA断裂水平。(I、J) 彗星实验检测基因组稳定性(cGAS缺失)。在敲除cGAS的NSFs中观察到DNA损伤显著增加。两组间差异的显著性通过Mann–Whitney U检验确定。
3 四个氨基酸决定裸鼹鼠cGAS的功能反转
为揭示裸鼹鼠cGAS特有的分子机制,研究者构建了人–鼠嵌合cGAS突变体,以定位关键结构域。结果发现,当人源C端替代裸鼹鼠C端时,其促进HR的能力消失;而当保留裸鼹鼠C端时,功能得以保留。进一步将C端分为三个亚结构域,结果显示C3区段(氨基酸位点444–554)是促进HR修复的关键模块。序列比对揭示该区域存在16个裸鼹鼠特有残基。
当这16个残基被替换成人类对应序列时,其促进HR的作用被完全消除;相反,将16个裸鼹鼠特征残基引入人类cGAS则消除了其抑制作用。进一步分析确定其中4个关键残基(S463D、E511K、Y527L、T530K)至少部分决定了这种功能逆转。当这4个位点突变后,裸鼹鼠cGAS失去了增强HR的能力,而在人类cGAS中引入对应突变(D431S、K479E、L495Y、K498T)则消除了其对HR的抑制效应。相应地,裸鼹鼠cGAS突变体无法再促进RAD51募集,而人类cGAS突变体则反转了其原本的抑制效应。这一四残基调控机制在小鼠中同样保守。
此外,这些突变并不影响cGAS的DNA感应功能,说明其在DNA修复中的作用与免疫功能相互独立。系统发育分析显示,这四个氨基酸位点的变化在裸鼹鼠相关谱系中逐步进化形成,而在其他啮齿类及长寿哺乳动物中均未出现。仅灰松鼠(Sciurus carolinensis)与盲鼹鼠(Spalax galili)在对应位置拥有相同残基(Y503与T606),与裸鼹鼠cGAS的Y527与T530相对应。
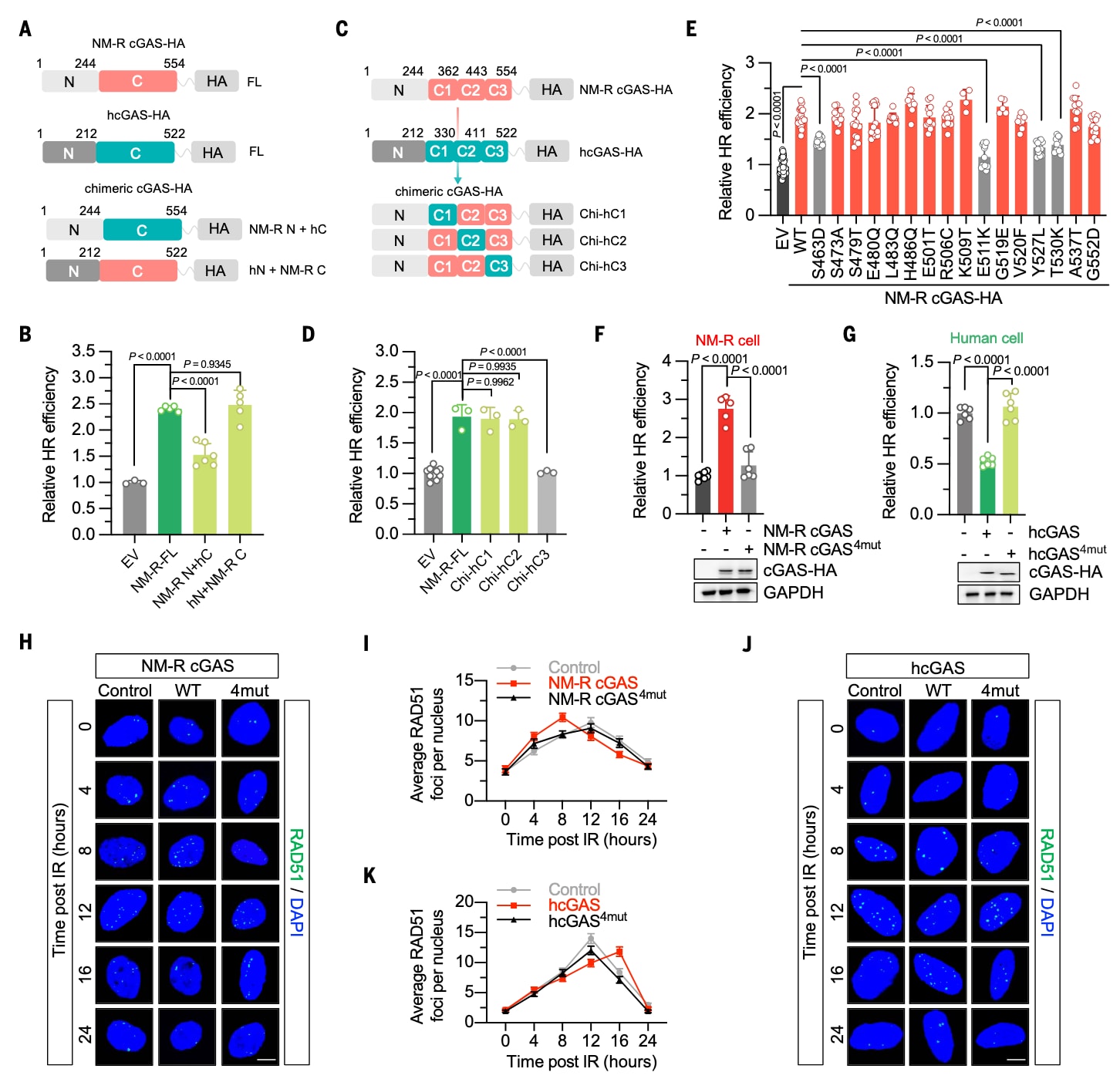
图2|四个氨基酸决定裸鼹鼠cGAS增强同源重组修复(HR)的能力 (A) 构建示意图。展示了带有HA标签的裸鼹鼠全长cGAS(NM-R cGAS-FL)、人类cGAS以及两者的嵌合构建体结构示意。(B) 嵌合cGAS对HR修复效率的影响。在裸鼹鼠成纤维细胞(NSFs)中过表达不同嵌合形式的cGAS,评估其对HR修复效率的影响。结果表明,含有人源C端的嵌合体失去增强HR的作用,而保留裸鼹鼠C端的构建体保持促进功能。显著性分析采用单因素方差分析(ANOVA)并经Tukey多重比较校正。(C) C端亚结构域替换示意图。将裸鼹鼠cGAS的C端分为三个亚结构域(C1–C3),并逐一以人类cGAS的相应序列替换。(D) 不同C端亚结构域替换对HR修复的影响。实验显示,仅当C3区段(位点444–554)保持裸鼹鼠序列时,HR修复效率显著提升。统计分析同样使用单因素方差分析(ANOVA)与Tukey校正。(E) 16个位点单残基突变的功能筛查。将裸鼹鼠cGAS的16个特征性残基分别替换为人类cGAS对应氨基酸后,检测其对HR修复效率的影响。氨基酸单字母缩写:A(丙氨酸)、C(半胱氨酸)、D(天冬氨酸)、E(谷氨酸)、F(苯丙氨酸)、G(甘氨酸)、H(组氨酸)、I(异亮氨酸)、K(赖氨酸)、L(亮氨酸)、M(甲硫氨酸)、N(天冬酰胺)、P(脯氨酸)、Q(谷酰胺)、R(精氨酸)、S(丝氨酸)、T(苏氨酸)、V(缬氨酸)、W(色氨酸)、Y(酪氨酸)。细胞共转染线性化HR报告系统、DsRed报告质粒及相应cGAS突变体表达载体,转染48小时后通过流式细胞仪(FACS)分析。显著性检验采用ANOVA与Dunnett’s T3多重比较。(F) 裸鼹鼠cGAS的四残基突变体显著削弱HR修复增强作用。在NSFs中过表达野生型(WT)或含四个关键突变(S463D+E511K+Y527L+T530K)的NM-R cGAS后,测得其HR修复能力下降。蛋白表达经Western blot验证。(G) 人类cGAS的四残基突变体失去HR抑制功能。在人源永生化皮肤成纤维细胞(HCA2-hTERT)中过表达野生型或四残基突变体(D431S+K479E+L495Y+K498T)的cGAS后,结果显示突变体逆转了原本的抑制效应。统计检验采用ANOVA与Dunnett’s多重比较。(H–K) RAD51灶形成动力学分析。在NSFs与HCA2-hTERT中分别过表达裸鼹鼠[(H)、(I)]或人类[(J)、(K)] cGAS的野生型或四残基突变体,观察辐照(8 Gy X射线)后不同时间点的RAD51荧光灶形成与消退过程(比例尺:10 μm)。结果显示,野生型NM-R cGAS促进RAD51快速募集与修复,而四残基突变体则明显减弱这一效应;在人类cGAS中,四残基突变体则消除了其对RAD51募集的抑制。误差线:在(B)、(D)–(G)中表示标准差(SD),在(I)、(K)中表示标准误差(SEM)。
4 裸鼹鼠cGAS的延长染色质结合由四个氨基酸变化介导
鉴于cGAS在DNA双链断裂(DSB)发生时会富集于损伤位点,研究者比较了DNA损伤后裸鼹鼠与人类cGAS在染色质上的保留时间差异。在接受8 Gy X射线照射的人胚肾细胞(HEK-293FT)中提取染色质后发现,两种cGAS均能结合至染色质,但裸鼹鼠cGAS在损伤后30分钟的结合强度明显高于人类cGAS(图3A)。
当在裸鼹鼠cGAS中引入四个关键氨基酸突变后,其从染色质上的解离显著加快;相反,人类cGAS的四残基突变体在DNA损伤30分钟后仍比野生型(WT)人类cGAS保留更多(图3A)。
DNA修复因子从染色质的分离通常依赖分离酶P97,该酶识别并提取带有K48连接型泛素链的底物蛋白。研究进一步比较了DNA损伤后人类与裸鼹鼠cGAS的泛素化状态。结果显示,电离辐照(IR)后人类cGAS的泛素化水平升高,而裸鼹鼠cGAS未出现此变化,并且这一差异由四个氨基酸突变决定(图3B)。
相应地,在照射15分钟后,裸鼹鼠cGAS与P97的结合显著下降,而人类cGAS与P97的相互作用则增强(图3C)。与之对应的是,裸鼹鼠cGAS四残基突变体表现出更强的cGAS–P97相互作用,而人类cGAS四残基突变体则消除了辐照后相互作用增强的现象。定量结果如下:NM-R cGAS,0.49 ± 0.28;NM-R cGAS⁴ᵐᵘᵗ,1.43 ± 0.13;人类cGAS,1.77 ± 0.30;hcGAS⁴ᵐᵘᵗ,0.56 ± 0.21(图3C)。在小鼠cGAS及其四残基突变体中进行的染色质结合、泛素化与P97互作实验也得到了一致结果(图S4)。
此外,既往研究表明DNA损伤可增强人类cGAS与E3泛素连接酶TRIM41的相互作用。为验证其作用差异,研究者检测了TRIM41对人类与裸鼹鼠cGAS泛素化的影响。共免疫沉淀(co-IP)实验表明,TRIM41的过表达使人类cGAS的泛素化水平提升约1.53倍,而对裸鼹鼠cGAS的泛素化几乎无影响(图3D)。进一步实验发现,在人类细胞中敲低TRIM41会降低cGAS的泛素化水平,而TRIM41过表达则促进cGAS与P97的相互作用(图3E、F)。
图3|DNA损伤后裸鼹鼠cGAS在染色质上的结合时间更长 (A) DNA损伤后cGAS染色质结合分析。在人胚肾细胞(HEK-293FT)中分别转染野生型(WT)或四残基突变体(4mut)的人类与裸鼹鼠cGAS,第二天以8 Gy X射线照射诱导DNA损伤。结果显示,裸鼹鼠cGAS在损伤后仍强烈结合于染色质,而其4mut突变体迅速解离;人类cGAS则相反,突变体在染色质上的保留更持久。灰度图显示了相对于组蛋白H3的cGAS-HA染色质结合量。(B) K48连接型泛素化水平分析。在HEK-293FT细胞中检测DNA损伤后15分钟内野生型或4mut人类/裸鼹鼠cGAS的K48型泛素化水平。结果表明,人类cGAS在照射后泛素化显著增强,而裸鼹鼠cGAS变化不明显,突变体的引入可逆转此差异。(C) cGAS与分离酶P97的相互作用。在照射15分钟后检测HEK-293FT细胞中P97与野生型或4mut人类/裸鼹鼠cGAS的结合。结果显示,裸鼹鼠cGAS与P97的结合减弱,而人类cGAS与P97的相互作用增强;突变后则呈相反趋势。(D) TRIM41过表达对cGAS泛素化的影响。在人源HEK-293FT细胞中过表达E3泛素连接酶TRIM41后,人类cGAS的K48型泛素化水平上升约1.5倍,而裸鼹鼠cGAS几乎未受影响。(E) TRIM41敲低对人类cGAS泛素化的影响。在HeLa细胞中敲低TRIM41后,DNA损伤15分钟时检测到cGAS的K48型泛素化水平明显下降。(F) TRIM41过表达增强人类cGAS与P97的结合。在HEK-293FT细胞中过表达TRIM41后,检测到P97–cGAS相互作用显著增强。以上所有实验均独立重复三次,定量结果以**平均值 ± 标准差(SD)**表示,并在图中各面板中展示。
5 裸鼹鼠cGAS通过FANCI介导的RAD50募集促进同源重组修复(HR)
由于裸鼹鼠cGAS在DNA损伤后能在染色质上长时间保持结合,研究者推测其可能充当DNA修复因子招募的平台。为验证这一假设,利用共免疫沉淀(co-IP)结合质谱分析,对经辐照处理的裸鼹鼠皮肤成纤维细胞提取物进行检测,结果鉴定出cGAS可能的结合靶蛋白——包括FANCI、RAD50和PARP1(图4A)。
在人类细胞中,cGAS已知通过干扰PARP1–Timeless复合物的形成而抑制HR修复。然而,在裸鼹鼠细胞中,未检测到PARP1与Timeless之间的相互作用(图S5A、B),且DNA损伤也未增强cGAS与PAR或PARP1的结合(图S5C、D)。因此,裸鼹鼠cGAS的HR促进作用并非通过调控PARP1–Timeless通路实现。
进一步的co-IP实验表明,带有GFP标签的裸鼹鼠cGAS可与带有HA标签的裸鼹鼠FANCI结合(图4B);由于缺乏针对裸鼹鼠蛋白的商业抗体,研究者在人体成纤维细胞中验证了该相互作用(图4C)。此外,裸鼹鼠RAD50与cGAS的结合亦通过外源与内源co-IP得到确认(图4D、E)。
RAD50是典型的DNA双链断裂修复因子,而FANCI主要参与链间交联修复。激光微照射实验显示,裸鼹鼠FANCI可被招募至激光诱导的DSB位点(图4F)。带标签的FANCI与RAD50共表达后可形成复合物,电离辐照(IR)刺激后二者的结合增强约1.84倍(图4G、H),体外co-IP实验亦证实其直接结合(图S6A)。这些结果提示FANCI与RAD50协同调控DSB修复,并可能受cGAS调控。
在人类细胞中同样观察到FANCI–RAD50的相互作用(图S6B),表明该复合物在进化上具有保守性。进一步mapping实验表明:FANCI的701–750与851–900位点以及RAD50的1–98位点是二者相互作用的关键区域(图S6C–H)。当删除这些区域(FANCIΔ851–900、RAD50Δ2–98)后,二者结合显著减弱(图S7A、B)。共过表达野生型FANCI与RAD50能协同增强HR修复,但当使用缺乏结合能力的突变体时,这一作用完全消失(图4I、J,图S7C、D)。因此,FANCI–RAD50复合物的形成对HR修复至关重要。
裸鼹鼠cGAS可增强FANCI与RAD50的结合,而人类cGAS则起相反作用(图4K,图S7E)。将16个或4个氨基酸位点替换后,裸鼹鼠cGAS对FANCI–RAD50相互作用的促进作用被削弱;而在人类与小鼠cGAS中引入相应突变,则转而促进二者结合(图4L、M,图S7F–H)。近邻连接分析(PLA)进一步证实,裸鼹鼠cGAS增强而人类cGAS破坏FANCI–RAD50结合,这种功能差异正是由四个关键氨基酸残基介导的(图4N、O)。
由于FANCI与RAD50之间存在直接结合(图S6A),裸鼹鼠cGAS在染色质上长期驻留可能提高二者相遇的概率,从而增强复合物形成并促进HR修复。相反,人类cGAS的短暂染色质结合可能导致FANCI与RAD50从染色质上被移除,表现出竞争性抑制作用。确实,当使用P97抑制剂CB-5083阻断其染色质脱离时,人类cGAS对FANCI–RAD50的抑制效应被消除,说明cGAS的染色质结合状态是其调控作用的关键(图4P)。
尽管人类cGAS的四残基突变体增强了FANCI–RAD50结合,但并未提升HR修复效率(图2G),因为其仍保持对Timeless–PARP1复合物的抑制作用(图S7I)。因此,这种负向调控效应可能抵消了突变体的正向影响。
RAD50结合DNA是DNA损伤响应中最早的事件之一,也是HR修复的关键步骤。染色质分级实验显示,裸鼹鼠FANCI能促进RAD50在DNA损伤后装载到染色质上(图4Q)。同时,过表达裸鼹鼠cGAS也能增强RAD50的染色质募集(图4R);而当FANCI被敲低时,这一效应完全消失(图4S),表明FANCI是裸鼹鼠cGAS介导RAD50招募所必需的。
进一步的免疫荧光实验显示,在接受辐照的裸鼹鼠细胞中,过表达裸鼹鼠cGAS能显著加速RAD51的招募,但在FANCI或RAD50被敲低的细胞中,这一促进作用消失(图4T、U,图S7J)。
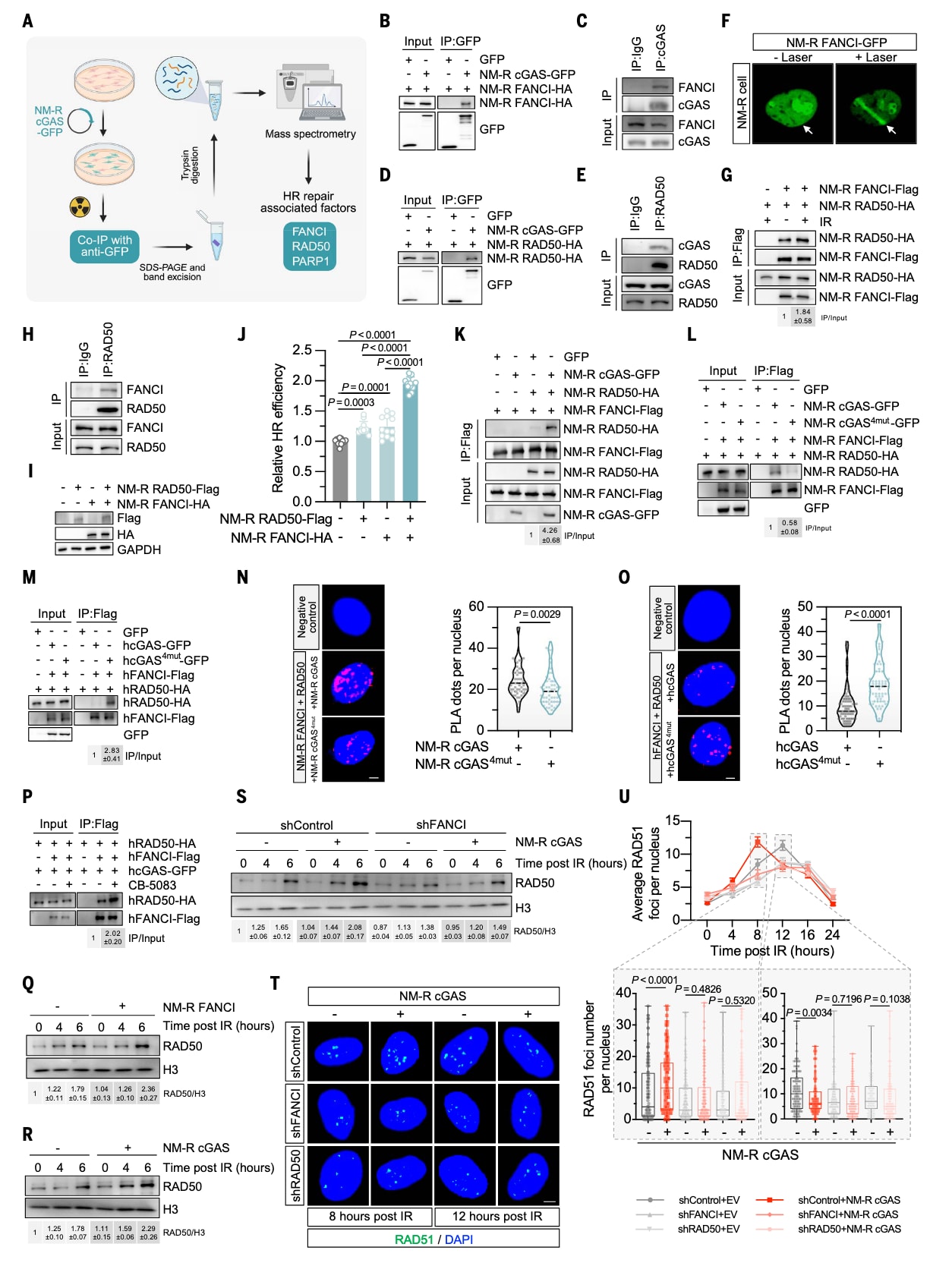
图4|裸鼹鼠cGAS通过增强FANCI–RAD50相互作用促进RAD50募集与同源重组修复 (A) 实验设计示意图。为鉴定与裸鼹鼠cGAS(NM-R cGAS)相互作用的同源重组(HR)修复相关因子,在裸鼹鼠成纤维细胞(NSFs)中转染带有GFP标签的NM-R cGAS,接受8 Gy X射线照射15分钟后进行共免疫沉淀(co-IP)及质谱分析(图由BioRender.com绘制)。(B、C) cGAS与FANCI的蛋白互作验证。在(B)中于NSFs中过表达NM-R cGAS与FANCI,进行co-IP验证;在(C)中使用人源成纤维细胞(HCA2-hTERT)检测内源性cGAS–FANCI相互作用。(D、E) cGAS与RAD50的蛋白互作验证。在(D)中于NSFs中过表达NM-R cGAS与RAD50进行co-IP检测;在(E)中进一步验证其在NSFs中的内源性结合。(F) FANCI在DNA损伤位点的招募。利用激光微照射诱导NSFs中DNA损伤,观察带GFP标签的NM-R FANCI在损伤位点的富集情况(箭头所示为照射位置)。(G、H) FANCI与RAD50的互作分析。在(G)中,NSFs在是否经8 Gy X射线处理后均进行co-IP检测,结果显示辐照增强FANCI–RAD50结合约1.8倍;(H)为其内源性交互验证。(I、J) FANCI与RAD50对HR修复效率的影响。在NSFs中过表达NM-R FANCI、RAD50或两者共表达,并使用空载体平衡转染量。(I)中通过Western blot验证蛋白表达,(J)中显示相对HR修复效率。结果表明共表达FANCI与RAD50显著增强HR修复。误差线表示标准差(SD)。(K) NM-R cGAS过表达增强FANCI–RAD50相互作用。在HEK-293FT细胞中过表达NM-R cGAS后,检测到FANCI与RAD50的结合增强。(L) NM-R cGAS四残基突变体对FANCI–RAD50互作的影响。突变后促进效应显著减弱。(M) 人类cGAS四残基突变体对FANCI–RAD50互作的影响。在人类cGAS中引入对应突变可增强FANCI–RAD50结合,与NM-R cGAS的变化方向相反。(N、O) PLA近邻连接实验分析。在HeLa细胞中分别检测过表达NM-R (N)与人类(O) cGAS及其突变体对FANCI–RAD50相互作用的影响(比例尺:5 μm)。结果表明,裸鼹鼠cGAS促进而人类cGAS破坏FANCI–RAD50结合,这种功能差异由四个关键氨基酸决定。(P) P97抑制剂对人类cGAS调控FANCI–RAD50相互作用的影响。使用10 μM P97抑制剂CB-5083处理HEK-293FT细胞4小时,结果显示该处理消除了人类cGAS对FANCI–RAD50结合的抑制效应,提示cGAS的染色质结合状态在调控中至关重要。(Q–S) RAD50在DNA损伤后染色质招募的变化。(Q) 过表达NM-R FANCI促进RAD50装载到染色质上;(R) 过表达NM-R cGAS同样增强RAD50染色质募集;(S) 在敲低FANCI的细胞中,这种由cGAS介导的促进作用完全消失。HEK-293FT细胞经8 Gy X射线照射后于4与6小时收集样本。(T、U) RAD51募集分析。在FANCI或RAD50缺失的NSFs中,经8 Gy X射线照射后于不同时间点检测RAD51的聚集情况。结果表明,NM-R cGAS在正常细胞中促进RAD51募集,但在缺乏FANCI或RAD50的背景下失效(比例尺:5 μm)。显著性采用Mann–Whitney U检验。
6 四个氨基酸突变调控细胞衰老、组织老化与寿命延长
虽然DNA修复能力受损会加速衰老,但增强DNA修复是否能延缓衰老仍不明确。研究者比较了裸鼹鼠与人类cGAS及其四残基突变体对应激诱导的早衰(SIPS)的影响。为排除cGAS经典DNA感应功能的干扰,构建了酶学失活型突变体(裸鼹鼠cGAS^E257A/D259A,简称cGAS^E/D;人类cGAS^E225A/D227A;小鼠cGAS^E211A/D213A),并在此基础上分别加入四残基突变。
在裸鼹鼠细胞中过表达cGAS^E/D可显著抑制SIPS,β-半乳糖苷酶染色及衰老标志物p21表达均明显下降;但引入四残基突变后,这种抑制作用完全消失(图5A–C)。相反,在人源成纤维细胞(HCA2-hTERT)中过表达人类或小鼠cGAS^E/D会增加衰老细胞比例与p21表达,而四残基突变则逆转该效应(图5D–F,图S8A–C)。结果表明,cGAS在不同物种中具有相反的衰老调控功能,而四个关键氨基酸决定了这一分化。
进一步利用软琼脂实验与小鼠异种移植模型评估其在肿瘤形成中的作用。结果显示,人类cGAS促进细胞转化,而引入裸鼹鼠样突变后转化作用被抑制;相反,裸鼹鼠cGAS抑制肿瘤发生,但突变后丧失该效应(图S8D–K)。
鉴于裸鼹鼠cGAS能长时间结合染色质并可能作为修复因子招募平台,研究者进一步探究其是否能在其他模式生物中延缓衰老。果蝇由于缺乏天然cGAS,因此被选作模型。实验构建了稳定表达裸鼹鼠cGAS^E/D、人类cGAS^E/D及其四残基突变体的转基因果蝇。RNA测序结果显示,裸鼹鼠cGAS^E/D与人类cGAS^4mut,E/D组的老年果蝇转录组接近年轻组,提示其可“重塑”衰老转录状态(图S8L)。
利用AgingClock基因集(包含33个随年龄下降的基因)进行富集分析(GSEA),发现裸鼹鼠cGAS^E/D组老年果蝇中AgingClock基因表达上升,而引入突变后该效应消失;人类cGAS^E/D则表现相反,突变后得以恢复(图5G)。此外,在老年果蝇肠道中,裸鼹鼠cGAS^E/D促进年轻型基因上调、衰老型基因下调,而人类cGAS^E/D则呈相反趋势(图S8M–N)。
老年果蝇常表现出免疫过度激活。GO分析显示,人类cGAS^E/D与裸鼹鼠cGAS^4mut,E/D组免疫相关基因表达升高,而裸鼹鼠cGAS^E/D未出现该现象(图S8O–P)。进一步检测肠道功能发现:过表达裸鼹鼠cGAS^E/D显著减少老年果蝇肠细胞增殖与肠干细胞(ISC)活性,而四残基突变体失去此保护作用;人类cGAS^E/D则加剧肠细胞增殖,突变后恢复正常(图5H–K)。
Smurf实验显示,裸鼹鼠cGAS^E/D可降低年龄相关肠道通透性,而人类cGAS^E/D则增加,突变同样能逆转(图S8Q)。在感染实验中,裸鼹鼠cGAS^E/D过表达的果蝇对肠球菌与粘质沙雷菌更具抵抗力并延长寿命,而人类cGAS^E/D缩短寿命,四残基突变则恢复耐受(图S8R–S)。
行为分析显示,裸鼹鼠cGAS^E/D改善老年果蝇的攀爬能力,而人类cGAS^E/D削弱之,突变后两者均被逆转(图S9A–B)。此外,裸鼹鼠cGAS^E/D减缓产卵能力衰退,而突变体无效;人类cGAS^4mut,E/D相比cGAS^E/D也表现出延迟的生殖下降(图S9C)。寿命分析结果表明,裸鼹鼠cGAS^E/D可显著延长果蝇的平均与最大寿命,而四残基突变消除了此效果;人类cGAS^E/D缩短寿命,突变体则恢复正常(图5L–M,图S9D),该趋势在不同环境温度下均保持一致(图S9E–H)。
在小鼠实验中,将携带裸鼹鼠cGAS^E/D、其突变体或对照载体的腺相关病毒(AAV)注入17个月龄小鼠尾静脉(图5N)。两个月后测定健康状况,结果显示:裸鼹鼠cGAS^E/D组体弱指数显著降低,而突变体组无改善(图5O)。肉眼观察可见,注射裸鼹鼠cGAS^E/D的小鼠皮肤病变较轻、毛发变白程度降低,但突变体失去此效果(图5P–Q,图S10A)。
进一步检测发现,裸鼹鼠cGAS^E/D组血清免疫球蛋白G(IgG)与白细胞介素-6(IL-6)水平显著低于突变组与对照组(图5R–S)。在肝脏中,裸鼹鼠cGAS^E/D减少衰老细胞累积与γH2AX标志物表达,而突变体失去此效应(图5T–W)。类似的抗衰老效应也出现在肾脏与肠道(图S10B–E),且伴随血清肌酐下降,提示肾功能改善(图S10F)。
综上,研究揭示:裸鼹鼠cGAS的四个关键氨基酸残基在调控细胞衰老、组织老化及寿命延长中具有决定性作用。这种通过DNA修复增强实现的抗衰老机制,为理解长寿进化与衰老干预提供了重要分子依据。
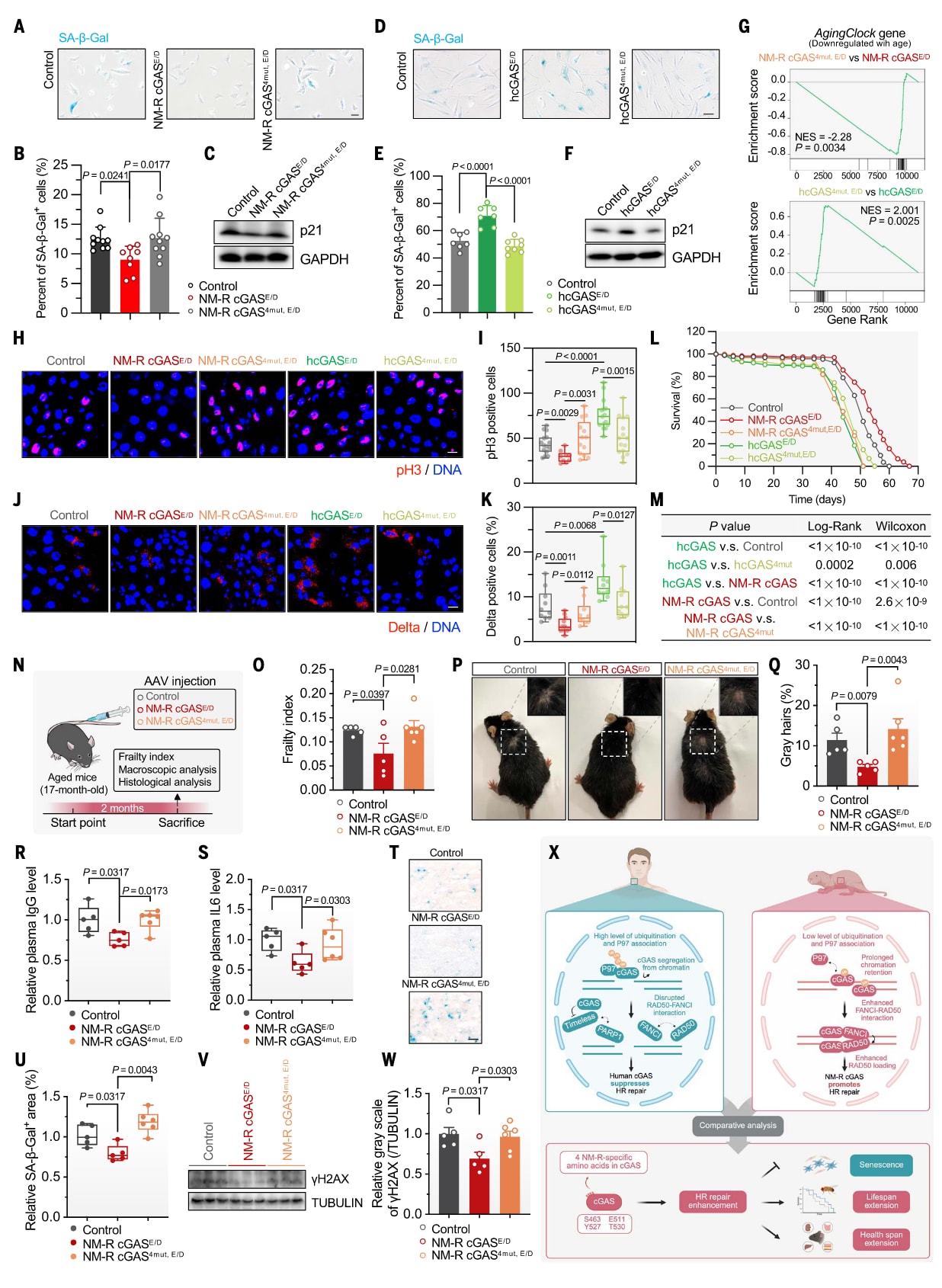
图5|四个氨基酸赋予裸鼹鼠cGAS更强的抗细胞衰老、延缓组织老化与延长寿命的能力 (A–C) 裸鼹鼠cGAS抑制早衰(SIPS)的效应。在裸鼹鼠成纤维细胞(NSFs)中过表达催化失活型NM-R cGAS(E257A/D259A,简称E/D)及其四残基突变体后,进行8 Gy X射线照射诱导SIPS。通过SA–β–gal染色[(A)、(B)]与p21蛋白水平(C)评估细胞衰老。结果显示,野生型NM-R cGAS显著抑制细胞衰老,而突变体失去此效应(比例尺:100 μm)。(D–F) 人类cGAS促进细胞衰老,突变后逆转。在人源成纤维细胞(HCA2-hTERT)中敲低内源hcGAS后,分别过表达催化失活型人类cGAS(E225A/D227A)及其四残基突变体,并以6 Gy X射线诱导SIPS。SA–β–gal染色[(D)、(E)]与p21表达(F)结果显示,人类cGAS促进衰老,而突变体逆转该现象(比例尺:200 μm)。统计分析采用单因素方差分析(ANOVA)与Tukey多重比较检验。(G) Drosophila肠道中AgingClock基因表达的GSEA富集分析。结果显示,裸鼹鼠cGAS^E/D显著上调AgingClock基因集(NES为标准化富集分值),而四残基突变后该效应消失。(H–K) 老年果蝇肠道的细胞增殖与干细胞活性分析。利用pH3免疫染色[(H)、(I)]与Notch配体Delta染色[(J)、(K)]检测老年果蝇肠道细胞增殖。结果显示,裸鼹鼠cGAS^E/D显著减少肠细胞分裂与干细胞活性,而突变体失去保护作用(比例尺:10 μm)。(L、M) 果蝇寿命分析。在29°C下,分别表达催化失活型人类/裸鼹鼠cGAS及其突变体的转基因果蝇进行寿命测定。结果显示,裸鼹鼠cGAS^E/D显著延长果蝇的平均与最大寿命,而突变体失去该效应;人类cGAS^E/D缩短寿命,突变体则恢复正常。显著性检验采用log-rank与Gehan–Breslow–Wilcoxon双检验。(N) 小鼠AAV注射实验设计示意图(使用NIAID NIH BioArt素材绘制)。在17月龄小鼠尾静脉注入携带裸鼹鼠cGAS^E/D、其四残基突变体或对照载体的AAV病毒。(O) 小鼠体弱指数(Frailty Index)分析。注射两个月后测量健康状况,结果显示:裸鼹鼠cGAS^E/D组体弱指数显著降低,而突变体无改善。(P、Q) 小鼠皮毛与皮肤状态。注射裸鼹鼠cGAS^E/D的老年小鼠毛发变白较轻、皮肤更健康,灰毛比例定量分析如(Q)所示(每只小鼠计数100根毛发)。(R、S) 血浆中IgG与IL-6水平检测(ELISA)。结果显示,裸鼹鼠cGAS^E/D组IgG与IL-6水平均显著低于突变体与对照组,提示全身炎症反应减轻。(T、U) 肝脏组织衰老检测。SA–β–gal染色(T)及定量分析(U)表明,裸鼹鼠cGAS^E/D显著减少肝脏衰老细胞数量(比例尺:50 μm)。(V、W) γH2AX水平分析。Western blot结果显示,裸鼹鼠cGAS^E/D组肝脏DNA双链断裂(DSBs)积累更少。显著性分析采用Mann–Whitney U检验。(X) 人类与裸鼹鼠cGAS功能机制模型图(由BioRender.com绘制)。在人类细胞中,DNA损伤诱导cGAS泛素化增加,从而增强P97介导的染色质解离,并破坏FANCI–RAD50复合物。同时,人类cGAS还削弱PARP1–Timeless相互作用,最终延迟RAD51募集并抑制HR修复。而在裸鼹鼠中,不存在PARP1–Timeless相互作用。cGAS的四个氨基酸变化使其泛素化水平降低、染色质结合更稳定,并增强与P97、FANCI–RAD50的协同作用,从而促进RAD50装载及RAD51募集,显著提升HR修复能力。这四个氨基酸的差异削弱了裸鼹鼠cGAS的抑制功能,使其获得更强的抗衰老潜力,延长果蝇寿命、减缓小鼠组织老化并改善生理健康。
7 讨论
裸鼹鼠的极端长寿被认为源于多种影响不同生物过程的适应性进化(45),包括细胞间通讯(46)、细胞周期停滞(47)、应激反应(48)、表观遗传调控(24)以及蛋白稳态维持(49),这些过程与衰老标志高度相关。然而,至今尚不清楚究竟是哪种分子机制使裸鼹鼠能够避免基因组不稳定。
研究表明,DNA双链断裂(DSB)修复能力与癌症发病率和年龄呈负相关(9, 22, 50–53)。在对18种寿命差异显著的啮齿动物的比较研究中发现,DSB修复效率与物种的最大寿命密切相关,其中长寿基因Sirt6在进化过程中可能受到正向选择(22)。值得注意的是,从长寿动物河狸中克隆得到的SIRT6酶活性显著高于短寿小鼠(22)。另有研究指出,关键HR修复因子BRCA1在裸鼹鼠中同样经历了正向选择(23)。然而,对于参与DSB修复的负调控因子是否也受到进化选择,一直缺乏关注。
DNA传感器cGAS在人体(20)和小鼠(fig. S1C)中被证实可抑制HR修复。该研究首次揭示,裸鼹鼠cGAS在HR修复中具有相反的功能,提示在这一长寿啮齿动物中,cGAS基因同样可能经历了正向进化。
RAD50是MRN复合物(MRE11–RAD50–NBS1)的关键组分,负责连接断裂的DNA末端及姐妹染色单体,以促进HR修复(54);而FANCI则是Fanconi贫血(FA)途径的重要成员,主要负责在链间交联修复中招募下游因子(14)。该研究发现,裸鼹鼠FANCI与RAD50之间的相互作用在应激条件下显著增强,且裸鼹鼠与人类cGAS对FANCI–RAD50复合物形成的影响方向相反。
与小鼠或人类细胞相比,裸鼹鼠细胞的生长速度更慢(55),这意味着其细胞内的DNA双链断裂可能在较长时间内得不到修复,从而引发无菌性炎症与衰老进程。在这种背景下,cGAS介导的FANCI–RAD50特异性调控或成为裸鼹鼠维持高效HR修复的重要机制。
研究进一步表明,裸鼹鼠cGAS与P97之间的相互作用减弱,使得cGAS能够更长时间地滞留在染色质上。这种延长的结合不仅促进了FANCI–RAD50的稳定复合,也可能为其他下游修复复合体的组装提供支架平台。此外,裸鼹鼠中cGAS功能的优化并非唯一促进DSB修复的机制,该研究还观察到诸如PARP1–Timeless相互作用的缺失等额外调节模式(图S5A、B)。
综合来看,该研究揭示了裸鼹鼠在进化过程中通过激活DNA修复功能以实现长寿的分子基础(图5X)。研究者确定了裸鼹鼠cGAS中四个关键氨基酸残基,正是它们赋予该蛋白增强HR修复的能力。研究结果支持“高效DNA修复可延缓衰老”的观点,并提示以cGAS为靶点调控DNA修复,有望成为促进健康寿命延长的潜在干预策略。