JMC 2025 | 利用噬菌体展示与冷冻电镜技术高通量鉴定并表征结合LptDE的双环肽
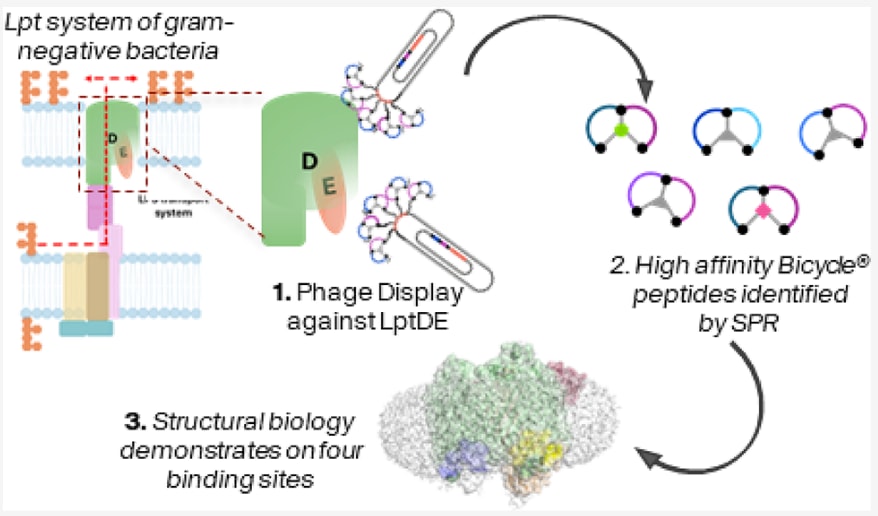
今天介绍的研究发表于 Journal of Medicinal Chemistry ,题为《利用噬菌体展示与冷冻电镜技术高通量鉴定并表征结合LptDE的双环肽》。革兰氏阴性菌的脂多糖转运系统(Lpt系统)维持其外膜完整性,其中LptDE复合物负责将脂多糖递送至外膜外层,是细菌生存必需的关键膜蛋白。由于LptDE外露于细胞表面且无需穿透膜即可被抑制,它成为新型抗菌药物开发的理想靶点。然而,目前尚无针对大肠杆菌或志贺菌LptDE的直接抑制剂。
该研究利用Bicycle噬菌体展示平台,结合表面等离子体共振(SPR)与冷冻电镜(cryo-EM),系统筛选并解析了志贺菌LptDE的双环肽结合模式。研究在LptD的β桶结构上发现了四个独立表位,并获得八个高分辨率复合物结构,揭示了保守的W×(D/EE)结合模体与特异性疏水作用。该研究建立了一种快速、可扩展的流程,用于膜蛋白靶点的高通量配体筛选与结构验证,为未来针对LptDE的抑制剂开发提供了结构基础与新策略。

获取详情及资源:
0 摘要
革兰氏阴性菌中的脂多糖转运系统(Lpt系统)在维持细菌外膜(OM)不对称结构的完整性方面起着关键作用。脂多糖的生物合成系统在大多数革兰氏阴性菌中都是必需的,其中LptDE复合物负责将脂多糖递送至外膜的外层。由于LptDE是一个位于细胞外、对生长至关重要的膜蛋白,它成为无需穿透细胞即可进行抑制剂开发的理想靶点。然而,目前尚无针对大肠杆菌LptDE的直接抑制剂,而药物研发面临挑战,因为该膜蛋白缺乏传统意义上的活性位点。
该研究利用双环肽噬菌体展示平台,结合冷冻电子显微镜(cryo-EM)和表面等离子体共振技术,鉴定并解析了志贺菌(Shigella flexneri)LptDE(SfLptDE)的双环肽结合模式。在SfLptD的β桶结构上共发现了四个不同表位,每个表位均具有独特的双环肽结合模体。该研究提出了一种高效的流程,可用于快速筛选与优先评估针对LptDE的候选结合分子,为后续抑制剂的发现提供了新策略。
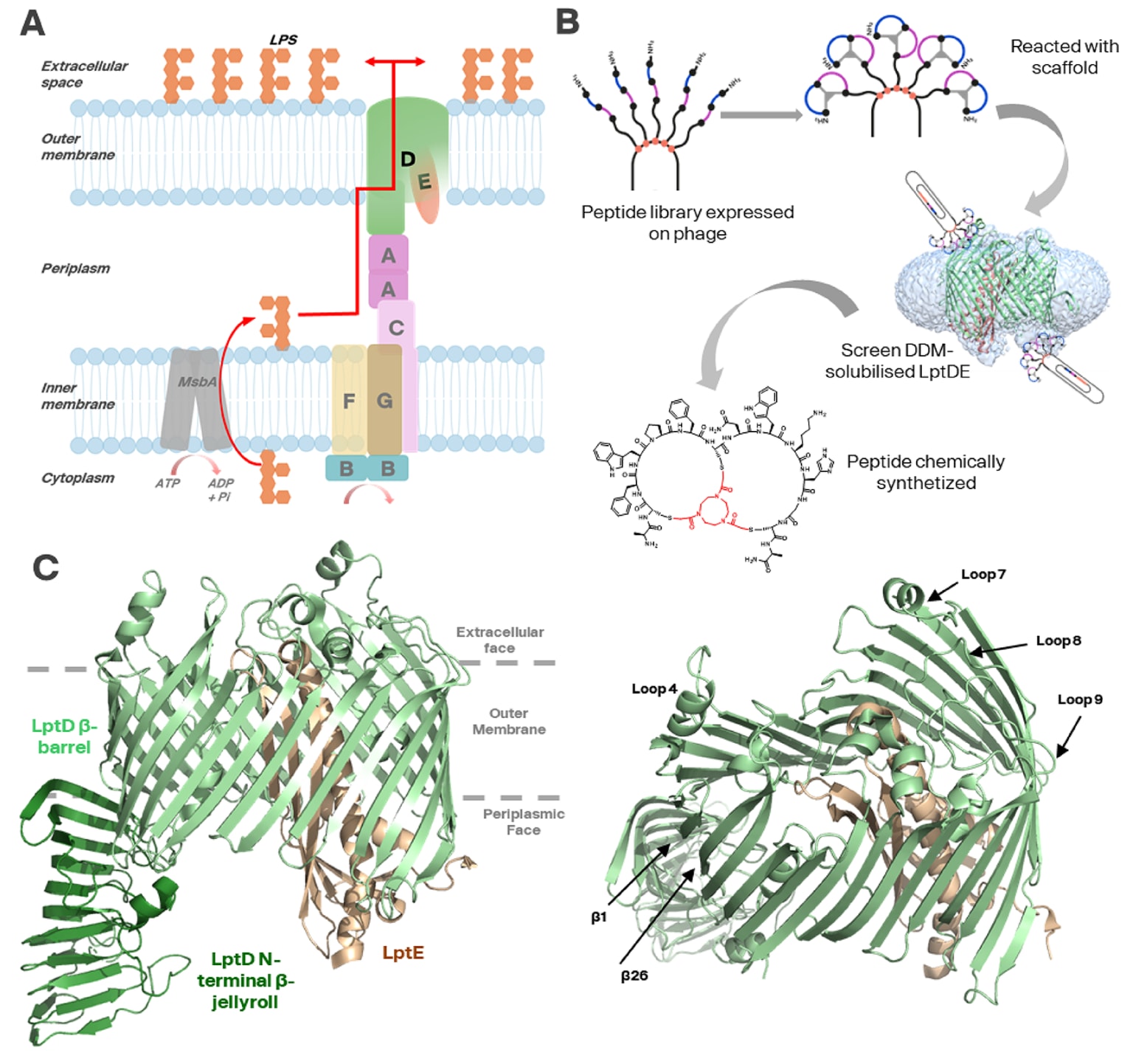
图1|针对志贺菌LptDE的双环肽噬菌体展示。 (A) LptDE是Lpt蛋白家族中最下游的成员,负责将脂多糖(LPS)从内膜运输至革兰氏阴性菌的外膜。该复合物由LptD与LptE组成。 (B) 基于靶点的Bicycle平台药物发现流程利用展示线性肽的噬菌体,这些肽随后通过化学支架(红色所示)环化形成双环肽,并以噬菌体形式展示。通过多轮亲和力筛选可富集与LptDE复合物结合的肽分子,之后进行化学合成(图示为化合物12,比例未按实际比例绘制)。 (C) SfLptDE复合物的晶体结构(PDB编号:4Q35),其N端β果冻卷结构域形成一个V形疏水沟,向周质侧延伸并与LptA接触。外膜相关的β桶由26条反平行β链(β1–β26)构成,这些β链在周质侧通过转角连接,而在细胞外侧则形成覆盖桶腔的延展环结构。LPS分子需通过这些环的重排才能到达外膜外层。LptE紧密嵌入LptD的β桶内部。
1 引言
多重耐药细菌正日益成为全球公共卫生的严重威胁,现有抗生素在治疗危险感染中的疗效正在不断削弱。这一趋势促使科研界迫切呼吁开发具有新化学骨架与创新作用机制的新型抗菌剂。对于革兰氏阴性菌而言,其独特的不对称外膜(outer membrane, OM)含有脂多糖(lipopolysaccharide, LPS)层,构成了阻止多数抗菌化合物渗透的屏障。七组分的LPS转运系统(LptA–G)通过将LPS从细胞质跨过周质层并运送至外膜外层,维持外膜的完整性。LPS首先由内膜复合物LptB₂FGC提取,经由由LptA组成的“蛋白桥”跨越周质层,最终抵达嵌入外膜的LptDE复合物(图1A)。
LptDE是一个由β桶蛋白LptD与脂蛋白LptE组成的“栓塞–桶状”(plug-and-barrel)复合体,跨越外膜,将LPS从周质空间转运至外膜外层。LptD包含两个结构域:位于周质中的N端β果冻卷结构域,以及嵌入外膜脂双层的β桶结构。LptE嵌入LptD的β桶内部,起到维持LptD装配与稳定性的作用(图1C)。LptDE复合物因其在大多数革兰氏阴性致病菌中的必需性及其外露的膜外位置,被认为是极具潜力的新型抗生素靶标。
然而,迄今为止针对LptDE的抑制剂筛选大多未获成功。Genentech曾开展大规模抗体筛选,仅获得无功能的抗-LptD抗体,推测原因为蛋白关键区域可及性不足。利用核糖体展示与噬菌体展示曾获得对淋病奈瑟菌(Neisseria gonorrhoeae)LptDE具有高亲和力的Pro-macrobodies,但并未展现抗菌活性。相比之下,最有前景的抗LptDE抗菌化合物是专用于铜绿假单胞菌的环肽Murepavadin(POL7080),其来源于天然抗菌肽protegrin-1。Murepavadin结合于该菌特有的LptD周质结构域。与细胞内靶点不同,直接作用于外膜蛋白(如LptDE)的分子无需穿透细胞膜,从而绕过了抗菌药物开发中的重要障碍。
该研究采用噬菌体展示技术发现针对这一困难膜靶点的双环肽(Bicycle分子)。Bicycle分子是一类双环肽,通过含三半胱氨酸的线性肽与三价支架发生反应形成两个环。环化产生的构象约束通常可提高靶标特异性与亲和力,同时增强抗蛋白酶降解的稳定性。Bicycle噬菌体展示平台提供了一种高通量、高效率的策略,用于筛选目标结合性环肽。这类化合物的化学空间类似于天然抗菌肽,但后者难以化学修饰,而Bicycle分子则可完全人工调控,便于药物优化。
冷冻电子显微镜(cryo-EM)能够在接近天然状态下以高分辨率解析膜蛋白结构,是结构基础药物设计的重要工具。尽管样品制备与成像通量仍具技术挑战,但近年来在制样、成像及数据处理技术的进步已使cryo-EM在复杂靶点上实现近原子分辨率。随着该技术的发展,针对历史上难以处理的膜蛋白靶点(如G蛋白偶联受体和离子通道)的分子设计已显著加速。
在此研究中,作者提出了一种结合Bicycle噬菌体展示输出、表面等离子体共振(SPR)检测及中等通量cryo-EM的综合方法,用于筛选并分类结合志贺菌(Shigella flexneri)LptDE(SfLptDE)的Bicycle分子。选择SfLptDE作为研究对象,是因为其与大肠杆菌LptDE具有高度序列同源性(两者仅有4个氨基酸差异),且便于制备。研究假设,针对SfLptD筛选出的Bicycle分子亦可结合E. coli LptD,因为两者序列差异中有2处位于膜埋藏区、1处在LptE的无序C端区域,不影响结合位点。
结果共发现多种能以优于1 μM亲和力结合SfLptDE的肽分子,并通过SPR竞争实验划分为多个独立结合类别。随后利用cryo-EM解析了其结合模式,获得了8个SfLptDE–Bicycle复合物的高分辨率结构,覆盖4个不同的结合位点。研究对cryo-EM数据采集策略进行了优化,可在24小时内生成多达3个分辨率足以确定结合表位的结构图,即使在小型膜蛋白体系中亦可实现。总体而言,该研究建立了一种稳健、高效的流程,用于快速识别、分类并定位LptDE结合分子,为膜蛋白靶向药物的发现提供了新思路。
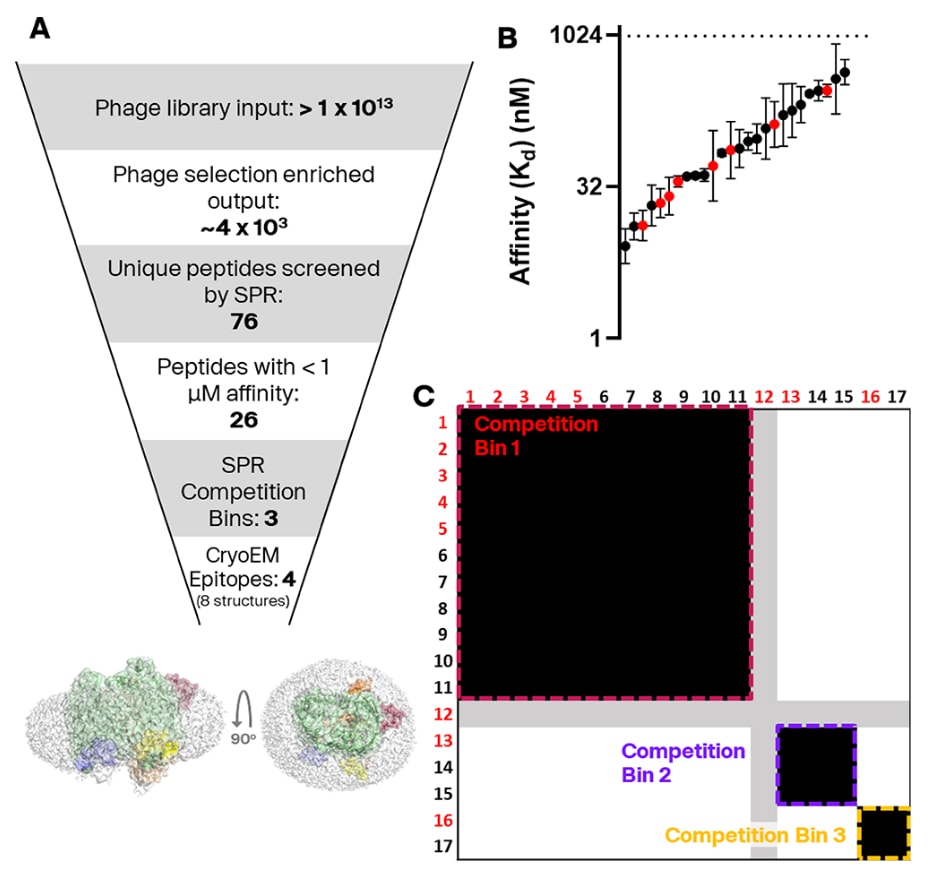
图2|筛选流程概览与SPR检测结果。 (A) 展示筛选流程的整体步骤示意。图中显示了双环肽分子与SfLptDE在四个不同表位上的结合情况(LptDE由胶束包围)。“竞争分组(competition bin)”与“表位(epitope)”的定义见正文说明。 (B) 通过表面等离子体共振(SPR)检测测定了76种肽分子与LptDE_FL的结合亲和力,其中26种肽的亲和力结果如图所示。其余50种肽被判定为无结合或非特异性结合。该实验中可可靠测得的最弱亲和力为1 μM。红色标示的化合物为后续用于冷冻电镜分析的样品。误差线表示至少两次独立生物重复的几何标准差。 (C) SPR竞争分组实验的热图视图。每一行/列代表一个肽分子的相互作用模式。肽–肽相互作用被分类为竞争(黑色)、非竞争(白色)或不确定(灰色)。每组相互作用的重复次数见补充图S3E。化合物12的相互作用结果无法明确分类,因为其未表现出与自身竞争的现象,而这一自竞争信号是关键的阳性对照,其代表性传感图见补充图S3D。
2 结果与讨论
2.1 噬菌体筛选鉴定结合分子
来源于志贺菌(Shigella flexneri)的LptDE(SfLptDE)在大肠杆菌BL21(DE3)细胞中共表达并纯化,随后重组入十二烷基-β-D-麦芽糖苷(DDM)胶束中。研究中使用了两种SfLptDE构建体:一种为全长形式(SfLptDE_FL),包含LptD的膜相关β桶与周质侧N端β果冻卷结构域(LptD残基25–784,LptE残基1–175);另一种为截短形式(SfLptDE_T),仅含LptD的β桶区段(LptD残基204–784,LptE残基19–193)。在大肠杆菌表达系统中,这两种构建体的LptD与LptE信号肽均被切除(见补充图S1)。
纯化后,两种SfLptDE复合物均经氨基反应型生物素酯化学标记,以便进行噬菌体展示筛选。生物素化的SfLptDE被用于筛选环化肽库,具体方法参考既有研究。噬菌体肽库由长度为11–17个氨基酸的肽组成,通过3个硫醚键与7种不同的小分子“支架”环化形成双环肽。筛选共进行四轮,每轮逐步降低SfLptDE浓度以提高筛选严格度。
研究开展了两组筛选实验:一组为针对全长SfLptDE_FL的小规模试验筛选,另一组为针对截短型SfLptDE_T的大规模筛选。选择截短构建体的目的是减少与LptD周质N端结构域结合的分子被富集,从而优先获得与外膜暴露区域结合的候选肽。结果表明,这两种蛋白形式均适用于噬菌体展示,利用高通量测序共鉴定出至少4000条独特肽序列,显示出多样的序列模体。
从两组筛选结果中,共选出76种Bicycle肽分子用于后续研究,筛选依据包括序列多样性、出现频率以及单克隆噬菌体在AlphaScreen实验中产生的阳性信号(见补充图S2)。这些肽随后采用固相肽合成(SPPS)进行化学合成,以开展生物物理特征分析(见图2)。
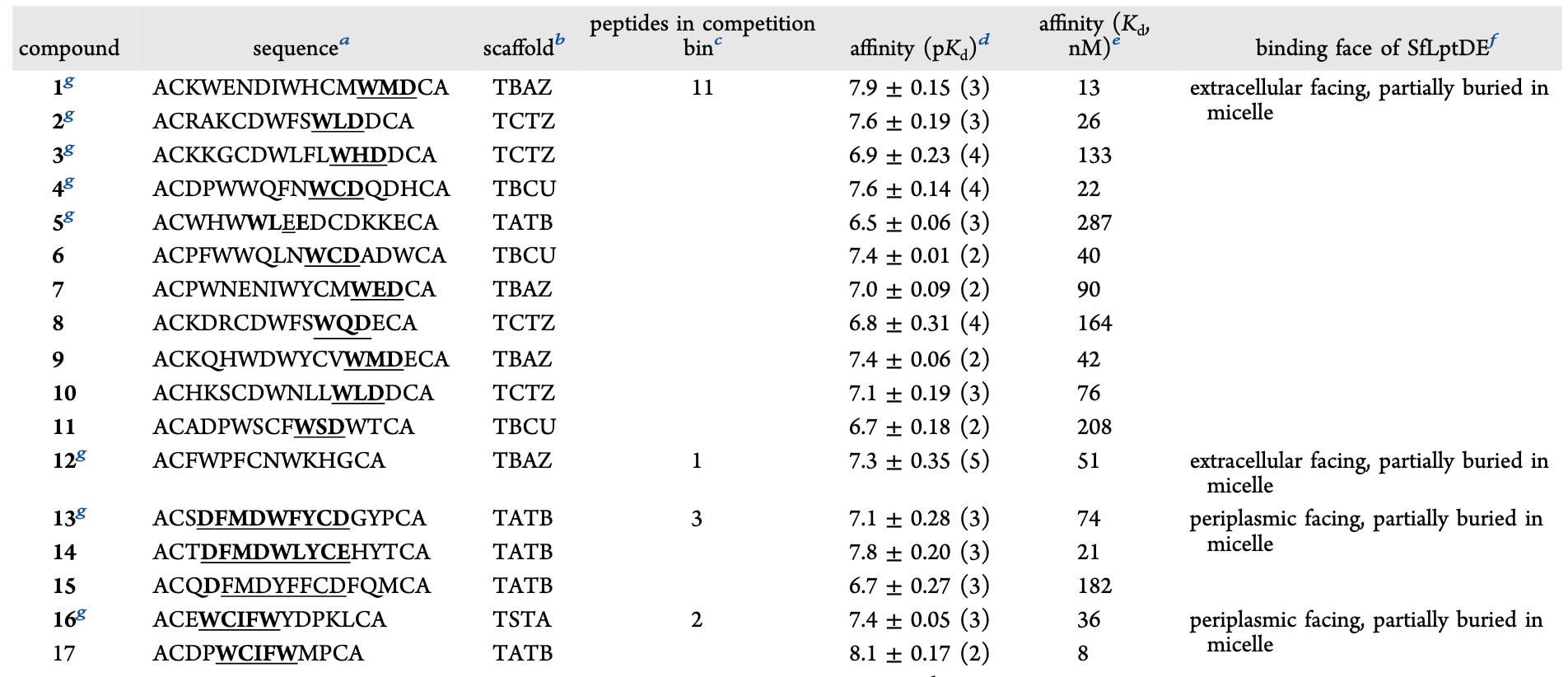
表1|该研究中鉴定出的与SfLptDE复合的双环肽分子特征
2.2 命中肽的生物物理分析
随后通过表面等离子体共振(SPR)实验测定双环肽分子与SfLptDE_FL的结合亲和力。SfLptDE在SPR检测中表现出高度可重复性,其响应曲线通常能良好拟合1:1结合模型,因此该方法被用于双环肽–蛋白相互作用的初步定量分析。在测试的76种双环肽中,有26种的结合亲和力优于1 μM,其中最高可达约10 nM(见图2C与表1)。
部分与SfLptDE结合的双环肽随后进入竞争分组实验(competition binning assay),采用SPR的A–B–A注射模式。该实验能够检测两种肽分子能否同时结合同一蛋白,从而判断它们是否竞争相同的结合位点(即同一“竞争分组”)。若两种肽竞争同一分组,通常意味着它们结合在蛋白的相似区域,但也可能受变构效应等其他因素影响。因此,文中“竞争分组(competition bin)”指通过SPR实验推测的结合位点,而“表位(epitope)”则指经结构生物学验证的结合区域。
对各竞争分组中肽序列的比对分析显示出保守的序列模体,表明肽的序列特征与其结合位置之间存在一定对应关系(见图2C与表1)。
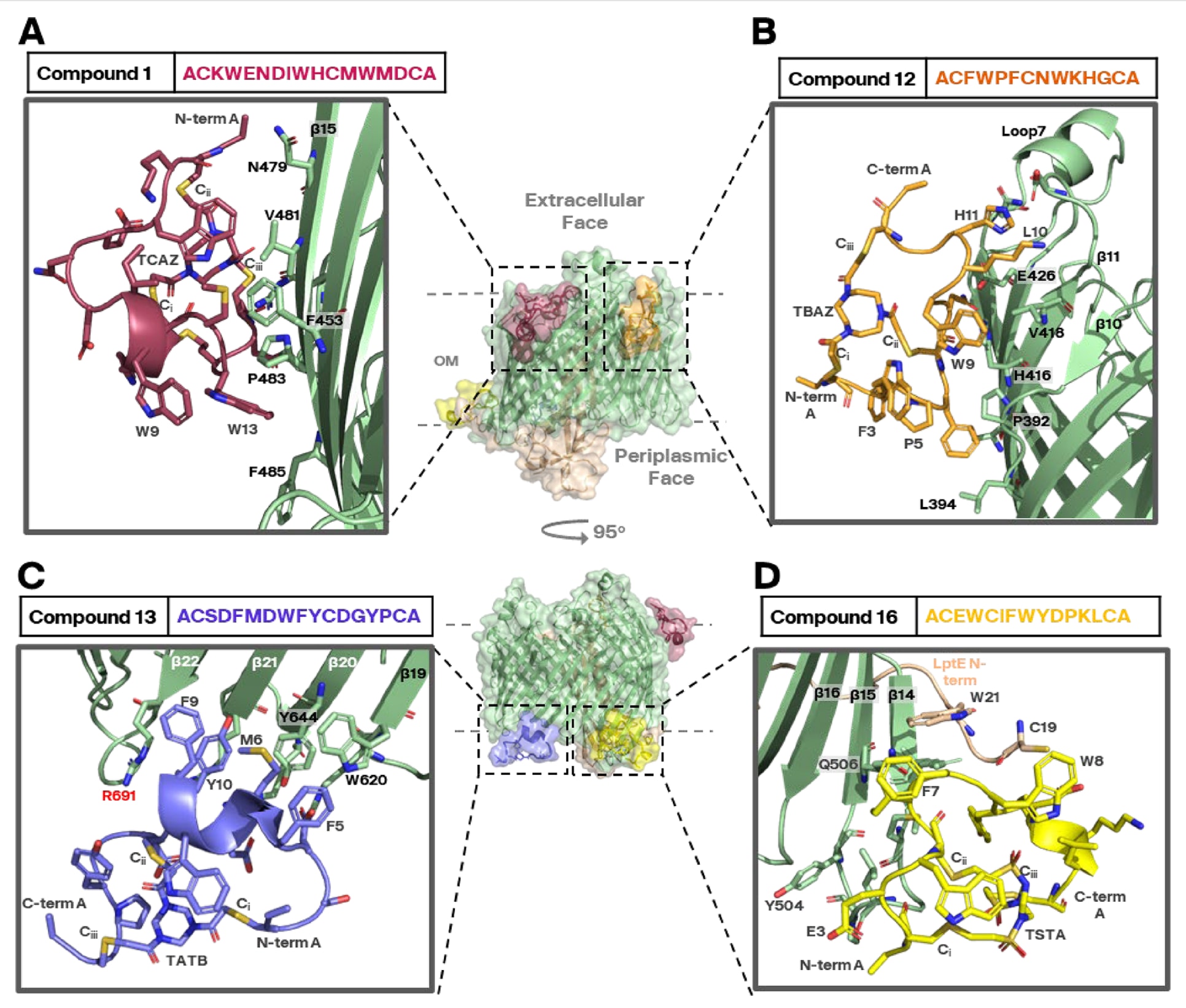
图3|双环肽分子与SfLptDE β桶复合物的结合模式。 冷冻电镜数据处理显示,四种双环肽分子(化合物1、12、13与16,PDB编号分别为9I92、9I97、9I98和9Q8N)分别结合于SfLptD蛋白的四个独立表位,位于蛋白的胞外侧(A、B)或周质侧(C、D)“外缘”区域。所有双环肽均具有两面:一面与蛋白结合,另一面与洗涤剂胶束接触。其与蛋白的相互作用主要通过疏水作用稳定,而带电残基通常朝向远离蛋白的一侧。整体来看,SfLptDE的主链在肽结合后与未结合状态(apo结构)相比几乎未发生明显构象变化(见补充图S5)。图中(C)红色标示的R691残基是E. coli LptD与SfLptD之间三个序列差异之一(见补充图S1A)。中心的复合叠加图仅用于示意说明,实际每个结构中SfLptDE仅与一个双环肽结合。所有双环肽结合位点的电子密度图见补充图S6。
2.3 冷冻电镜结构分析
从每个竞争分组中选取一个具有代表性的双环肽分子,与SfLptDE_FL形成复合物并通过冷冻电子显微镜(cryo-EM)进行结构解析。首个数据集(化合物12,图3B,PDB编号9I97)在24小时内共采集约1万帧图像,初步提取得到约120万个颗粒,最终利用约33万个颗粒重建出分辨率为2.48 Å的三维结构。该分辨率足以解析SfLptDE的β桶结构与双环肽分子(见图3A及补充图S4),结果显示双环肽结合在蛋白胞外侧β桶外缘的β片层11–13之间。虽然肽分子与蛋白形成了明确的肽–蛋白接触,但分子的大部分结构嵌入于洗涤剂胶束中,未引起桶结构的明显重排(见补充图S5)。
为评估数据采集效率,将提取的颗粒按采集时间分组(1小时、2小时、3小时)分别处理,分析获得亚3 Å分辨率所需的最小颗粒数。结果显示,在不影响最终分辨率的前提下,每个样品的数据采集可缩短至6–8小时,单次采集约3500帧。后续所有数据集均采用该策略,最终重建的分辨率范围为2.3–2.9 Å(见补充图S4及表S2)。这一优化使得每24小时可获得三套高分辨率结构。
在所有数据集中,双环肽分子均可在电子密度图中清晰构建,显示其在SfLptDE复合体上存在四个独立表位的结合模式。所有肽在各自表位上的结合方式相似,如化合物12所示,其一面与β桶外缘相互作用,另一面与洗涤剂胶束接触(见图3)。整体上,四个结合位置分别为:
(1)胞外侧β片层11–13(化合物12,PDB 9I97,图3B);
(2)胞外侧β片层14–16及外向环7(化合物1,PDB 9I92,图3A);
(3)胞质侧β片层15–16(化合物16,PDB 9Q8N,图3D);
(4)胞质侧β片层20–21(化合物13,PDB 9I98,图3C)。
在所有重建中,N端β果冻卷结构域的分辨率明显低于膜内β桶区。尽管通过聚焦分类与补充数据采集可部分改善,但由于双环肽不针对该区域,通常不再进行额外处理(见补充图S4E)。
其中,化合物16结合于SfLptD β桶胞质侧的β片层15–17,靠近LptE的N端区域,即其脂蛋白尾部锚定膜的位置。该复合物的重建结构显示额外密度信号,与LptE N端半胱氨酸延伸的脂尾相一致(见补充图S8)。此前的LptDE结构及其他配体复合结构均未观察到此处密度,因此推测双环肽的结合稳定了该区域。
化合物13的结合位点位于SfLptD胞质侧β片层20–22附近,包含关键残基Arg691(图3C中红色标示)。根据补充图S1,该位点在大肠杆菌LptD中为色氨酸,因此化合物13不太可能结合E. coli LptDE,因为在SfLptD中,化合物13的Asp12羟基与Arg691胍基形成的氢键是结合稳定的关键(见补充图S1与S7)。
为验证SPR竞争分组结果,又采集了来自竞争分组1的四个不同双环肽的数据集。该分组包含最多的独特序列。重建结果显示,这四个肽均结合于相同区域,且结合方式与化合物1高度一致(见图4A),表明SPR分组能够有效区分不同结合位置的分子。
在该表位中成像的五个分子(化合物1–5)具有一致的结合特征:其序列中存在保守的W×(D/EE)模体,与SfLptD的β桶形成稳定相互作用(见图4B)。其中的色氨酸残基嵌入由Pro483、Phe485、Tyr516和Pro518组成的疏水口袋内;Arg520与Tyr516通过氢键进一步稳定该模体,与D/EE残基的主链和侧链均形成接触(见补充图S7)。其他肽段,尤其是面向洗涤剂胶束的一侧,排列一致性较差(见图4与表1)。
突变实验进一步验证了该模体的重要性:将W残基突变为Ala完全消除肽的结合能力,而将Asp/Glu替换为Ala显著降低结合亲和力(见图4与补充表S3)。这说明尽管双环肽与SfLptDE的结合界面相对较小,其结合仍具有较高的序列特异性,高通量筛选过程并未仅筛出非特异性与胶束结合的分子。然而,当突变位于面向胶束的疏水环区时(如化合物1),结合同样被完全破坏(见图4与补充表S3),表明胶束环境在双环肽–SfLptDE结合的稳定性中也起到重要作用。
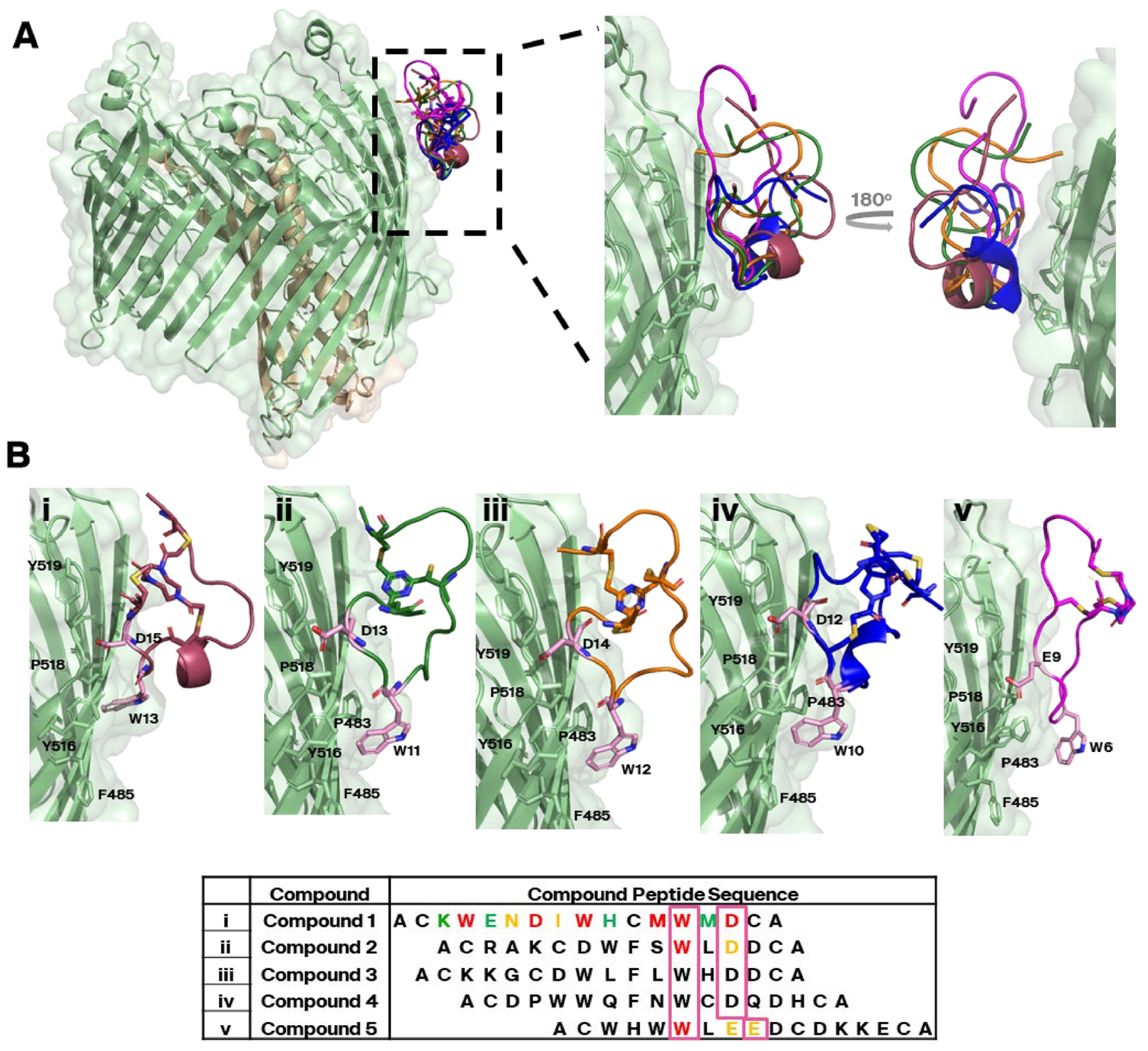
图4|同一竞争分组(竞争分组1)中的双环肽均结合于SfLptDE的相同位置(表位1)。 (A) 五种双环肽分子(化合物1–5,PDB编号分别为9I92、9I93、9I94、9I95和9I96)叠加图显示,这些属于竞争分组1的分子均结合于SfLptDE β桶胞外侧外缘的相同区域(表位1),具体位于β片层11–13之间。为便于观察,该图中去除了肽分子的化学支架部分。 (B i–v) 五种肽均通过保守的W×(D/EE)模体(粉色框)与蛋白形成接触。该相互作用主要由疏水侧链稳定,并辅以少量LptD桶结构的氢键作用(见补充图S7),其余分子部分对齐较差,显示结合的模体具有序列特异性,而非全分子一致性。在接触LptDE的模体之外,各肽的序列多样性较高。化合物1、2与5进行了丙氨酸扫描实验:绿色残基替换为丙氨酸后结合亲和力几乎不变或变化极小;黄色残基替换导致亲和力显著下降;红色残基替换则完全丧失与SfLptDE的结合能力;黑色残基未参与突变测试。各肽的结合亲和力数值见补充表S3。
3 结论
LptDE因其在革兰氏阴性菌外膜组装与脂多糖(LPS)转运中的关键作用,被认为是抗菌药物研发领域的重要新靶点。该研究旨在发现能够结合LptDE并潜在抑制LPS运输的分子,从而为无需细胞通透的抗菌剂开发提供思路。研究建立了一套用于鉴定与表征双环肽(Bicycle分子)结合SfLptDE不同表位的系统流程。
针对LptDE的药物发现面临多重挑战,其中包括缺乏可直接作用于该蛋白的化合物,以及缺少可用于功能验证的抑制实验。在候选分子的验证阶段,即便是针对LptDE这样至关重要的靶点,化合物活性可能过弱而难以在细胞生长实验中体现。通过敲除相关蛋白的化学基因组学策略可以增强细胞对抗菌化合物的敏感性,但即便在该研究中使用了外膜减弱的多种突变菌株,也未观察到对这些双环肽的敏感性。
在膜蛋白靶点的筛选中,最主要的困难之一是如何以生理相关的构象呈现目标蛋白。近年来通过将靶点嵌入洗涤剂胶束中实现了这一点。然而,洗涤剂的存在也带来了额外问题,即如何明确区分真正与蛋白结合的分子与仅与洗涤剂相互作用的分子。稳定的蛋白纯化流程、快速的数据采集与处理在该研究中发挥了关键作用,使得可以在较短时间内解析多个结合位点的多种配体复合结构。
目前唯一得到验证的LptDE抑制剂Murepavadin尚未获得其与LptDE复合物的结构信息,但耐药突变研究显示其结合位点位于铜绿假单胞菌LptDE的N端结构域。该结构域在假单胞菌与志贺菌间差异极大,而两者结构比较表明,Murepavadin耐药突变位点与该研究发现的四个结合表位相距较远。虽然该研究未考察所获分子是否能结合假单胞菌LptDE,但在对应表位上,志贺菌与假单胞菌蛋白存在一定的保守残基。
该研究主要识别到与洗涤剂界面结合的分子,而非直接导致功能抑制的肽,其原因尚不完全明确。一种可能是肠杆菌科LptDE(如大肠杆菌与志贺菌)的蛋白在缺乏假单胞菌特有结构域时,对抑制剂的“可成药性”较低。另一种可能是由于洗涤剂DDM在双环肽背面的较大接触界面产生偏好性结合,从而提升了与洗涤剂结合的亲和力。通过改进蛋白制备体系或可减少这种偏差,例如采用其他膜蛋白呈现策略,包括:在细胞中过表达LptDE进行整细胞筛选、使用外膜囊泡、脂质体、纳米盘或SMALPs系统等。这些方法在纯化与筛选方面仍具挑战,但值得进一步探索以优化筛选流程。
该研究在早期发现阶段引入了冷冻电镜技术,展示其在靶点范围内系统探索潜在配体化学空间的能力。共解析了8个双环肽与LptDE结合于4个表位的高分辨率结构,所有配体在一定程度上与洗涤剂胶束相互作用。高分辨率的结构使得能够清晰识别与洗涤剂结合的分子,从而在早期实现有效的筛选淘汰。鉴于目前学界与工业界对外膜靶点在原核与真核系统中均高度关注,这项工作提供了一种具有广泛适用性的策略,可帮助药物研发人员在早期阶段识别并排除非特异性洗涤剂结合分子,从而提升靶点导向药物发现的效率。