Nature 2025 | 结构快照解析μ-阿片受体中核苷酸释放的构象机制
今天介绍的这项工作来自 Nature。该论文围绕μ-阿片受体(MOR)如何以结构方式调控G蛋白核苷酸交换这一关键药理学问题展开,通过多种信号检测与放射性配体结合实验建立了“配体效能与GDP亲和力降低、GDP释放加速”之间的定量关联,并进一步结合单颗粒冷冻电镜与分子动力学模拟,在纳洛酮与洛哌丁胺条件下解析出覆盖激活早期的6种构象,其中包括4个GDP结合中间态(latent、engaged、unlatched、primed),系统捕捉到α5螺旋延伸、AHD逐步开启以及核苷酸结合口袋协调残基持续减少的连续重排路径。结构比较显示,拮抗剂或低效能配体倾向于将复合物滞留在GDP协调较完整、释放较慢的latent态,而高效能激动剂更易推动进入为AHD开启与GDP释放“预备”的engaged及其后续开放态,从而降低GDP再结合概率并提高核苷酸交换效率。总体而言,研究提出配体通过改变MOR-Gi复合物在非活化到latent再到engaged、unlatched与primed等构象之间的平衡来跨越GDP释放这一限速步骤,为解释配体效能差异提供了高分辨率的结构框架,并为GPCR功能选择性与更安全阿片类药物的理性设计提供了可推广的机制线索。

获取详情及资源:
0 摘要
μ-阿片受体(MOR)属于G蛋白偶联受体超家族,其通过促使Gα的α-螺旋结构域(AHD)打开来激活异源三聚体G蛋白,从而为GDP-GTP交换提供空间;其中,GDP释放被认为是该交换过程的限速步骤。基于药理学实验可见,激动剂的效能与GDP亲和力降低呈相关性,这种降低倾向于促进GTP交换;相对地,拮抗剂会提高GDP亲和力,从而抑制受体激活。围绕这一现象的结构基础,相关研究给出了MOR与纳洛酮或洛哌丁胺复合时的8个独特结构模型以及16张冷冻电镜密度图,由此捕捉到沿激活通路分布的多个中间构象。其中包含4种GDP结合态,展示了此前未描述的受体-G蛋白界面形式、AHD的空间排布,以及为实现GDP释放而在核苷酸结合口袋内发生的关键过渡。结构上,纳洛酮使MOR停滞在一种潜伏状态;而洛哌丁胺则推动体系进入一种结合状态,该状态在构型上已为AHD打开以及随后的GDP释放做好准备。分子动力学模拟对这些观察提供了支撑,并进一步指出,GDP结合的中间体以及AHD的构象选择是决定核苷酸交换速率的重要因素,从而为理解G蛋白激活机制与配体效能提供了结构与机理层面的线索,并对G蛋白偶联受体药理学具有更广泛的启示。
1 引言
在G蛋白偶联受体(GPCR)超家族中,μ-阿片受体(MOR)因其突出的临床重要性而尤为关键,并被广泛作为镇痛治疗的药物靶点。然而,靶向MOR的激动剂具有较高的滥用风险,且与潜在致命的不良反应相关,使得如何在药物开发中缓解这些问题成为当前策略的重点。用于治疗阿片类药物过量的调节剂已经存在,其中纳洛酮是临床应用最广泛的代表;它通常被视为拮抗剂,但既往研究也提示其可能具有低效能的部分激动活性。总体而言,部分激动剂被认为有望在维持疗效的同时获得更安全的治疗特征,优于传统MOR激动剂;不过,由于对于不同程度激动作用如何在细胞内被传递仍缺乏充分的机理认识,因此实现按需定制效能的配体设计仍受到限制。
阿片类激动剂通过与阿片受体结合来调控信号转导,而这类受体主要经由Gi/o型G蛋白传递信号。MOR介导的信号过程涉及其与G蛋白相互作用,推动G蛋白从非活化的GDP结合异源三聚体形式(Gαβγ)转变为活化状态,即由GTP取代GDP并促使复合物解离为GTP结合的Gα与Gβγ亚基。该过程的限速步骤是Gα亚基中GDP的释放,因为在静息条件下GDP被紧密结合。完成活化后,各亚基进一步作用于下游效应器,从而传播诸如腺苷酸环化酶抑制以及钾离子通道激活等信号。
在Gα亚基内部,核苷酸的结合、交换与GTP水解由Ras同源结构域(RHD)与α-螺旋结构域(AHD)界面处保守的结构基序所调控。RHD上负责协调核苷酸的关键元件包括β6-α5环中的TCAT基序,以及位于α1上的P环,后者与核苷酸磷酸基团结合;在AHD侧,Switch I与αE、αF共同提供额外的配位接触。已有观点提出,受体诱导的α5螺旋向受体内部的伸展、旋转与平移,可能破坏核苷酸结合稳定性,从而促进GDP释放。
在GPCR药理学中,配体通常依据其调节下游信号的方式及幅度进行分类,这一性质被称为效能:拮抗剂阻断激活,完全激动剂产生最大激活,部分激动剂引发低于最大值的响应,而超激动剂则可产生高于参考激动剂的超最大响应。因此,一个核心问题在于,不同水平的配体效能如何与受体-G蛋白复合物的构象状态相联系。尽管文献中提出了多种模型,激动剂激活异源三聚体G蛋白能力的差异,常被归因于GDP亲和力下降、GTP亲和力上升或二者共同作用。此前已有多项研究报道,GPCR激动剂的效能与其对应G蛋白的GDP亲和力降低之间存在直接相关性;此外,在MOR体系中的单分子Förster共振能量转移与双电子-电子共振实验也显示,配体效能与GDP释放速率相关,其机制可能是通过调节核苷酸结合的受体-G蛋白复合物状态之间的平衡来实现。总体而言,GDP结合态在G蛋白激活中具有关键作用,但迄今缺乏这些中间态的高分辨率结构快照,从而限制了对GPCR信号机理的深入理解。基于药理学实验、单颗粒冷冻电镜(cryoEM)研究与分子动力学(MD)模拟的综合证据表明,效能确与GDP释放速率相关,而该速率可通过稳定特定构象状态而被调控。
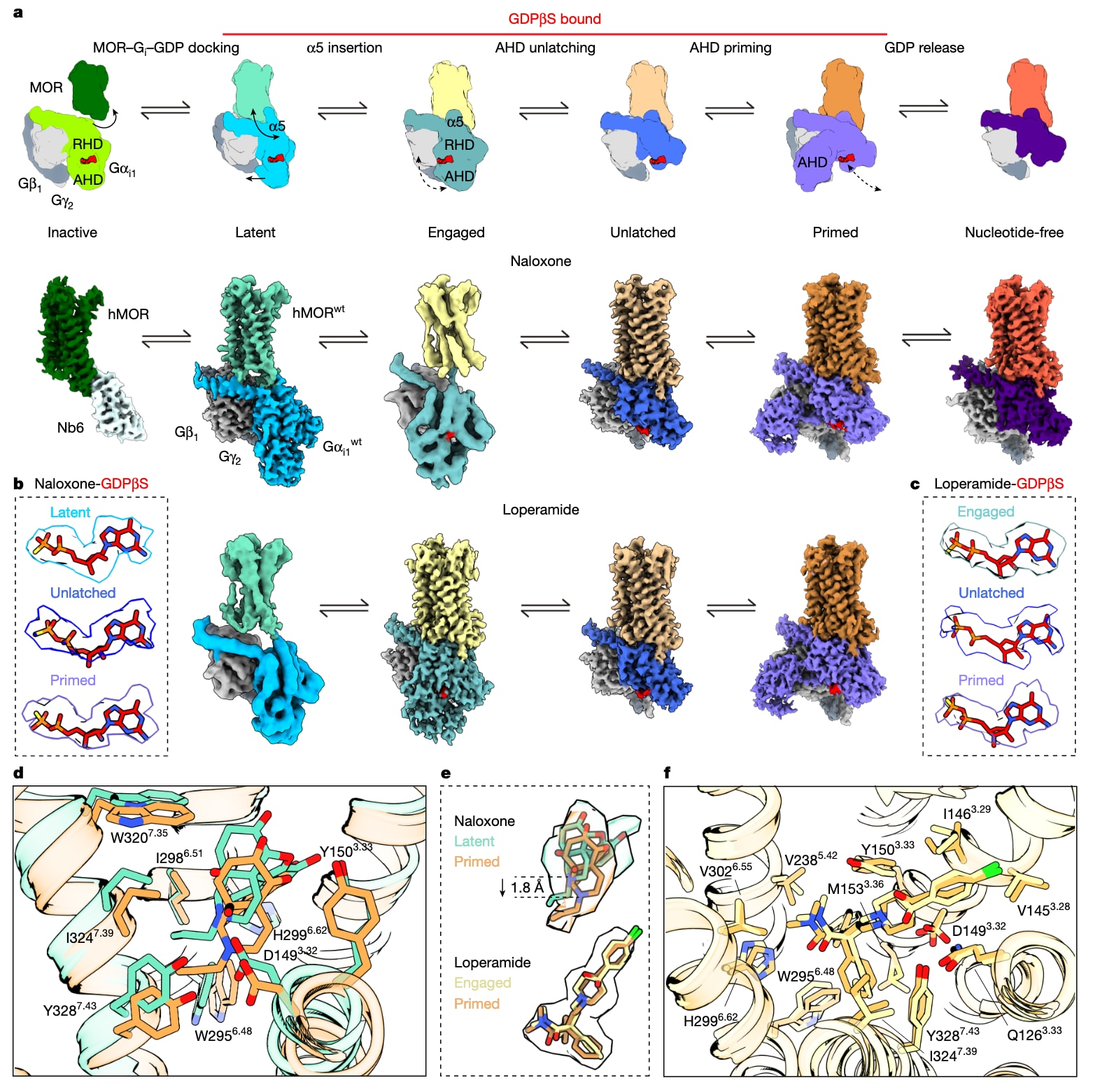
图1 | 展示了MOR在G蛋白激活通路早期阶段的结构快照。 a,展示MOR及MOR-Gαi1复合物在纳洛酮或洛哌丁胺存在下的cryoEM重构概览,共解析得到6种构象状态,并覆盖有无GDPβS的条件,依次为:(1)非活化,(2)latent,(3)engaged,(4)unlatched,(5)primed,(6)无核苷酸态。b,c,给出对应G蛋白结构中GDPβS的密度分布;b为纳洛酮体系下的latent、unlatched与primed;c为洛哌丁胺体系下的engaged、unlatched与primed。d,展示纳洛酮在正构结合口袋内于latent与primed两种状态之间的结合情况,并以棒状形式标出参与相互作用的残基。e,叠合展示正构口袋内配体的整体姿态变化:纳洛酮在latent与primed结构之间呈现“浅”与“深”的姿态差异,其中浅绿为latent,橙色为primed;洛哌丁胺在engaged与primed结构之间的叠合显示其姿态,其中黄色为engaged,橙色为primed。f,展示洛哌丁胺在正构结合口袋内于engaged与primed两种状态之间的结合情况,并以棒状形式标出参与相互作用的残基。wt表示野生型。
2 结果
2.1 GDP亲和力与MOR配体效能相关
为澄清纳洛酮效能表述上的矛盾,相关研究首先开展了体外信号实验,采用基于生物发光共振能量转移(BRET)的多类生物传感器,分别监测G蛋白异源三聚体状态(TRUPATH)、受体-G蛋白相互作用(RG-BRET)、ONE-GO工具集,并以cAMP生物传感器(GloSensor)作为下游信号读出。实验比较了反向激动剂阿维莫潘(alvimopan)、纳洛酮、部分激动剂mitragynine pseudoindoxyl、参考完全激动剂(D-Ala2,N-MePhe4,Gly-ol)-脑啡肽(DAMGO)以及超激动剂洛哌丁胺(loperamide,Lop)。结果显示,纳洛酮的效能位于阿维莫潘与mitragynine pseudoindoxyl之间,在所有检测体系中均明确呈现部分激动活性,幅度约为10%至30%。
为进一步从结合层面验证纳洛酮在MOR上的部分激动特征,研究进行了放射性配体结合实验,使用与纳洛酮结构相近的
在此基础上,研究进一步检验“配体效能与核苷酸结合亲和力相关”的假设,并在RG-BRET框架下建立了可读出核苷酸剂量依赖BRET变化的实验,将其命名为“nuc-BRET”,用BRET信号变化作为核苷酸亲和力的替代指标。为减少内源性核苷酸的竞争,研究用地高辛样去垢剂digitonin使表达MOR-Rluc8与Gαi1β1γ2-GFP2的细胞通透化,并用apyrase水解残余核苷酸,随后以不可水解的核苷酸类似物GDPβS与GTPγS进行剂量-反应实验。无配体(apo)条件下,MOR的GDPβS
为在细胞环境中进一步检验这一趋势,研究也在完整细胞中进行了实验,总体趋势相同,但阿维莫潘与纳洛酮引发的信号变化过小而难以解释,推测与内源性核苷酸竞争有关。细胞质中GDP与GTP的平均浓度分别约为50μM与500μM,提示两者持续存在竞争压力。为模拟该竞争,nuc-BRET实验在逐步提高GDPβS浓度(0μM,0.01μM,0.1μM,1μM)的条件下测定GTPγS
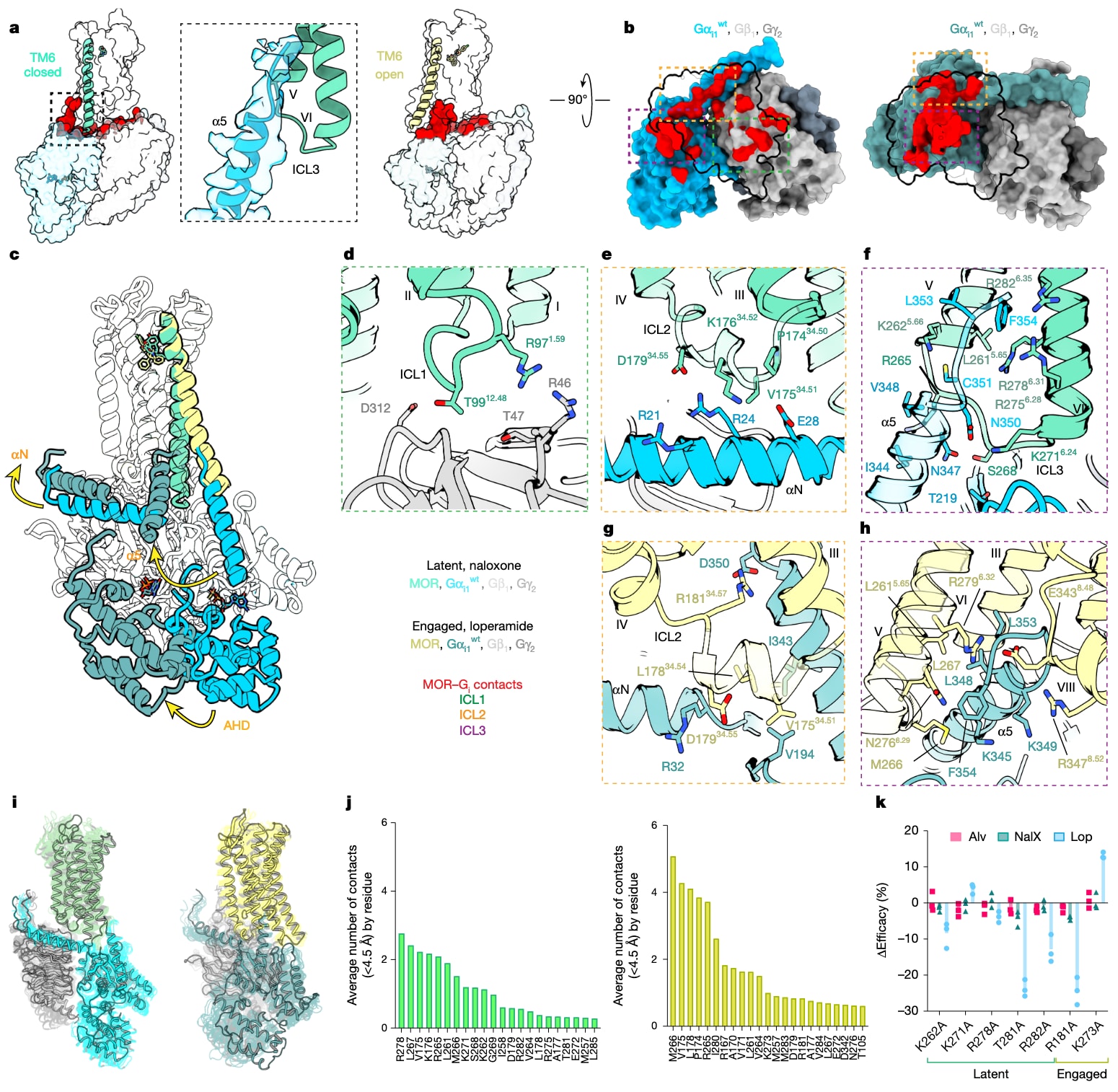
图2 | 展示了GDP结合的MOR-Gαi中间态结构比较:latent与engaged构象。 a,侧视结构模型对比latent与engaged两种状态下的MOR-Gαi1界面,插图强调Gαi1C端的有序密度及其与TM5、TM6和ICL3的结合;红色标示MOR与Gαi1β1γ2之间4Å以内的相互作用残基,突出通过各ICL介导的相互作用重排。b,俯视结构模型对比latent与engaged状态下的界面,并以黑色勾勒受体轮廓;彩色虚线框标示不同ICL界面(绿色ICL1,橙色ICL2,紫色ICL3)。c,以示意图对比两种状态下的MOR-Gαi1,并用箭头强调Gαi异源三聚体相对MOR发生的显著整体重排。d,latent状态下ICL1与Gβ1残基的相互作用。e,latent状态下ICL2与αN的相互作用。f,α5与TM5、TM6及ICL3的相互作用。g,engaged状态下ICL2与αN的相互作用。h,α5与TM6、TM7及H8的相互作用。i,latent与engaged状态的MD模拟示意,不透明模型为MD初始结构,透明结构为随机选取的后续帧。j,latent与engaged状态下MOR-Gαi1界面的MD统计,曲线表示各自状态在5条MD轨迹中超过1μs的平均接触数(距离阈值大于4.5Å)。k,针对两种状态各自特有残基( latent对比engaged)进行突变的RG-BRET实验,给出相对野生型构建体的归一化效能;柱状图为
2.2 MOR-G蛋白激活通路的结构快照
为从机理层面理解GDP亲和力如何被调控,相关研究进一步采用单颗粒冷冻电镜(cryoEM)实验获取结构证据。基于前述nuc-BRET结果,研究推测在不引入工程化突变的前提下,仅使用野生型受体与G蛋白序列,也有机会捕捉到GDP结合的MOR-Gi复合物中间态。为制备GDP结合样品,研究先获得无核苷酸的复合物,并分别与纳洛酮或洛哌丁胺结合,随后在冰上加入100μM的GDPβS孵育1h后再进行冷冻制样,将该流程称为“再结合GDP”(re-bound GDP)。该策略的设计依据来自GPCR药理学中一系列成熟的实验传统:由于GDP的释放与再结合被认为具有可逆性,许多研究会从无核苷酸复合物出发,再补加GDP,以探究GDP结合态GPCR-G蛋白复合物的作用与性质;这一思路在多种实验体系中均被沿用,包括放射性配体结合、
为进一步检验该流程得到的结论是否稳健,研究还设置了对照条件,即在纯化全过程始终加入GDP,作为“恒定GDP”(constant GDP)方案,并在后续分析中与“再结合GDP”策略进行比较。此外,研究评估了细胞水平的Mg
2.3 驱动GDP释放的构象变化
通过单颗粒冷冻电镜(cryoEM)流程,相关研究在MOR激活通路上解析得到6种构象状态。其中,来自相应样品的4种GDPβS结合态为早期G蛋白结合与激活提供了结构快照,并被命名为latent、engaged、unlatched与primed。沿着这四个状态可观察到一系列逐步推进的构象重排,其总体特征是参与协调GDP的残基数量逐渐减少,提示GDP结合口袋正在被系统性“松动”。为覆盖G蛋白偶联之前与核苷酸释放之后的关键参照点,研究同时解析了非活化MOR结构(3.4Å)以及无核苷酸的MOR-Gi结构(3.0Å)。围绕配体效能的机理讨论,全文重点对比了四个代表性结构:纳洛酮诱导的latent态(3.8Å),以及洛哌丁胺诱导的engaged态(3.2Å)、unlatched态(3.2Å)与primed态(2.9Å);并在这些结构基础上开展了大规模分子动力学(MD)模拟,用于检验界面与构象转变的稳定性与动力学特征。
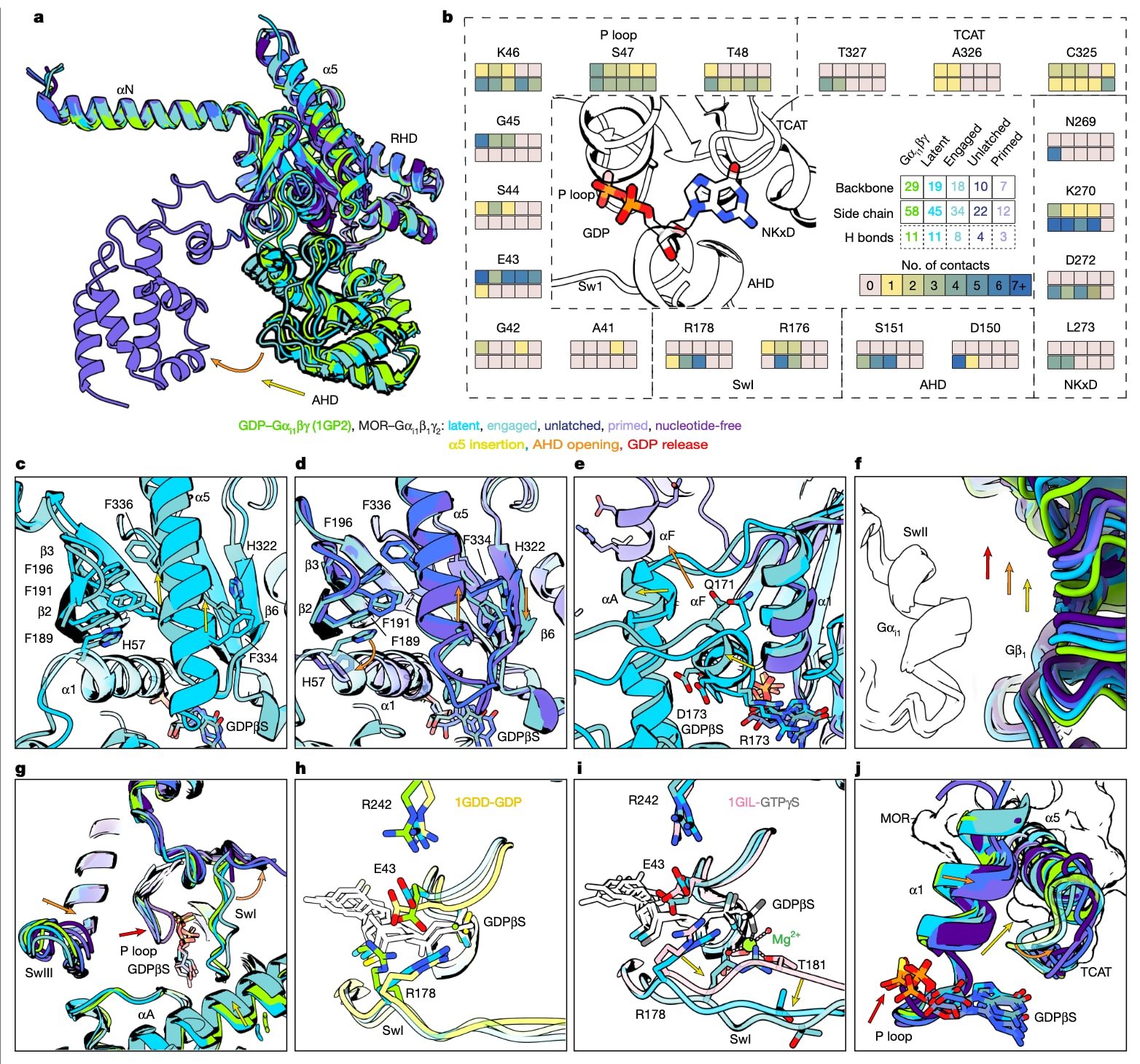
图3 | 展示了GDP释放前各构象状态下Gαi1及其核苷酸结合口袋的比较。 a,叠合展示受体游离的GDP-Gαi1β1γ2(PDB编号1GP2)与latent、engaged、unlatched、primed状态下的Gαi亚基结构。b,以接触图总结上述5种状态中Gαi残基与GDPβS的相互作用,接触由LigPlot+判定,并以颜色编码每个状态与残基的接触数量(浅红表示0次,深蓝表示每个残基与GDP超过7次相互作用);第一行实线框表示主链相互作用,第二行实线框表示侧链相互作用,第三行虚线框表示氢键,中心仅给出氢键总数。c,展示Gαi疏水核心的概览,包含α5螺旋、α1与α3,并强调从latent到engaged转变过程中的构象变化,尤其是α5向受体核心平移以及TCAT环的部分回撤。d,进一步展示疏水核心(包含α5、TCAT、α1与α3)在engaged、unlatched与primed三种状态间的变化,重点强调α5的平移与旋转导致TCAT环回撤以及β6的重排。e,展示GDP释放所需的αF转变与α1移动,并对比latent、engaged与primed状态。f,展示Gα与Gβ亚基之间相对夹角的连续变化趋势。g,对比不同状态下SwI、P环与SwIII。h,展示核苷酸结合口袋内E43与R178从GDP-Gαi1β1γ2(PDB编号1GP2)中闭合的“安全带”构象向latent状态重排的过程。i,展示engaged状态下R178向Mg
2.3.1 latent态MOR-Gi
在latent态中,非活化MOR与GDP结合的Gαi1β1γ2之间的初始“接触式”结合呈现出一个独特界面。与既往所有已解析的相关结构不同,该状态下G蛋白相对受体发生约60°的旋转,并通过全部三个细胞内环(ICLs)同时参与结合:ICL1与Gβ1形成接触,ICL2与αN螺旋相互作用并与R24关联,而在无核苷酸状态下对应的相互作用更接近R32;ICL3则与α5螺旋形成广泛相互作用。
G蛋白激活的早期关键步骤之一,被认为是打破Gα亚基内部由α5、β2-β3与α1共同形成的疏水核心,该过程发生在α5向受体核心延伸期间,并与GDP释放相联系。在latent态结构中,ICL3与多个构成或连接该疏水核心的基序存在接触,提示该界面可能直接参与“撬动”疏水核心。将latent态G蛋白与不含受体的GDP-Gαi1β1γ2结构(PDB:1GP2)进行对比可见多处关键差异:在latent态中,GαC端α5螺旋呈延伸状态且密度完整可辨,其末端4个残基伸入去垢剂胶束区域,并与跨膜螺旋TM5和TM6的胞质端发生接触。末端F354被TM6上的R275^6.28、R278^6.31、R282^6.35以及T281^6.34等残基环绕;其中,上标为GPCR的Ballesteros-Weinstein编号。这些残基对协调保守的DRY基序具有关键作用,暗示该相互作用可能承担调控功能。
latent态界面最复杂的部分位于ICL3与Gαi异源三聚体关键基序之间。ICL3与TM6向Gαi与Gβ1之间的裂隙伸入,该区域由R265、M266、S268、K271^6.24等残基构成,并与α5螺旋、αN螺旋及D261附近残基形成相互作用。MD模拟支持该latent态受体-G蛋白界面具有稳定性,其相互作用残基总数与无核苷酸状态相近,但平均接触数更少。为验证该结合模式的生理相关性,研究对latent态相对于engaged或“类活化”状态特有的界面残基进行了突变测试,多数突变体表现为配体效能降低,支持该状态在信号传导中具有实际作用。
在核苷酸结合口袋层面,latent态仍保留了不少与GDP相关的主链与侧链相互作用,涉及RHD中的E43-T48(P环)、K270、E272、L273(β5/αG)、C325-T327(TCAT环),以及AHD中的D150-S151(αD-αE环)、R176(αF)、R178(SwI)等。然而,整体相互作用数量明显下降,主链相互作用由29降至19,侧链相互作用由58降至45。该减少部分可由E43(P环)、D150(αD-αE环)与N269(NKxD)侧链接触的部分或完全丢失来解释;相对地,R176(αF)与R178(SwI)的侧链相互作用有所增加。
在受体游离条件下,Gβγ与Gαi1结合会促进E43与R178之间形成氢键,从而稳定一种类似“安全带”的构象,该构象遮挡核苷酸结合口袋并可能阻碍GDP释放。在latent态中,E43失去与R178的这一相互作用,而R178转而协调GDP的磷酸基团,这一模式与GDP结合的单体Gαi以及某些GDP结合的异源三聚体结构中的观察一致,并且该转变也被预测与后续GTP结合与水解的需求相关。与此同时,D150与核糖及鸟嘌呤基团之间的大部分非极性接触也发生丢失。综合这些重排,latent态结构呈现出一个可能的“成核点”,用于启动GDP释放并引发AHD的开启过程。与该推断一致,尽管部分MD轨迹与受体游离的GDP-Gαi1β1γ2较为接近,仍可观察到AHD短暂开启的事件,提示latent态已具备推动核苷酸交换的早期动力学倾向。
2.3.2 latent态MOR-Gi
在向engaged态转变时,G蛋白发生一系列显著的整体运动:α5螺旋延伸约14Å,并沿其最长轴旋转约60°,同时整体向MOR方向平移约6Å,且进一步向RHD方向下沉约3Å,由此形成与无核苷酸状态相同的受体-G蛋白界面。与此同时,MOR本身呈现出经典的受体活化特征,包括TM6外张,并伴随微开关(microswitch)基序的典型构象重排。这些变化与关键的G蛋白-核苷酸接触被破坏同步发生,并促使保守残基F336从由β2、β3与α1构成的疏水口袋中移位,从而改变稳定GDP结合的相互作用网络。
在engaged态中,AHD整体仍保持闭合构象,但相对RHD朝向αN方向旋转约13°,同时αA螺旋出现一定程度的拉直。核苷酸结合口袋内部,TCAT环表现为部分回撤,但C325与GDP之间的氢键仍保持。随着α5向受体方向平移,α5上的F334指向TCAT环,而β6上的H322仍指向Gαi的C端。与此同时,α1/P环向TCAT方向移动约1.2Å,并在后续构象状态中延续这一移动趋势。
与AHD旋转相伴,口袋内出现一组更具指示性的局部重排:位于αF上的R176侧链构象上抬约20°并朝向TCAT;位于αE上的D150失去与GDP残余的疏水接触,此时AHD中与GDP保持相互作用的残基仅剩S151。此外,Switch I(SwI)基序发生重新构型,其部分打开并向α1方向外展,导致R178近乎旋转90°。R178的胍基进入原本形成Mg
整体而言,与GDP的相互作用进一步减少,主链相互作用由19降至18,侧链相互作用由45降至34。鉴于SwI对GDP的协调对于维持AHD闭合构象至关重要,因此该短暂状态中观察到的重排很可能触发AHD开启的起始过程。与此一致,MD模拟显示其中3条轨迹出现AHD的短暂开启,另有1条轨迹呈现AHD的完全开启,支持engaged态已具备向核苷酸释放方向推进的动力学倾向。
2.3.3 unlatched态MOR-Gi
unlatched态呈现出高度动态的AHD构象,这一特征与既往报道的无核苷酸状态MOR-G蛋白结构相似。尽管核苷酸结合口袋周围发生了一系列构象变化,冷冻电镜密度仍能清晰指示GDP的存在。此状态下,F334与H322分别发生侧链构象翻转:F334转而指向Gαi的C端,而H322改为指向TCAT环。这一变化提示TCAT环正完成向完全回撤构象的最终过渡,并与AHD开启同步发生。与此同时,AHD与SwI中所有原本参与核苷酸协调的残基相互作用均消失,包括S151、R176与R178。此外,P环主链中S44与G45对GDP的相互作用也被解除。
由于协调作用显著丢失(主链相互作用由18降至10,侧链相互作用由34降至22,氢键由8降至4),GDP整体从其结合口袋向外移动约2Å。α1进一步朝向α5移动,并以极性相互作用替代了原先由被移位的β6-α5环所提供的疏水接触。α1末端两个残基I56与H57原本参与疏水核心的形成,在AHD开启后出现无序化。该螺旋继续向α5靠近,其位置与无核苷酸状态一致,并且Q52对TCAT环提供支撑。
另一个值得注意的现象出现在Gβ1上:T128-E130环在此前状态中密度解析度较差,而在unlatched态中则呈现清晰可辨的密度,并相对于latent与engaged态更为回撤,反映其对AHD开启的结构响应。MD模拟进一步显示,unlatched态具有最显著的AHD构象异质性,从完全开启到几乎完全遮挡均可出现。在AHD开启的轨迹中,可观察到鸟嘌呤基团的释放,而核苷酸仍可通过磷酸基团维持结合。该整体异质性可能解释了该cryoEM重构中AHD密度表现为无序的原因,同时也提示短暂的再遮挡状态可能限制GDP的进一步释放。
2.3.4 primed态MOR-Gi
在下一步转变中,开启的AHD趋于有序化,其连接区与AHD本体在结构上都能被清晰解析,并沿Gβ1亚基表面贴靠排列。该界面的关键相互作用来自Gβ1上的R134与R137,它们参与对AHD的定位与协调。与AHD闭合的“非活化”G蛋白状态(latent与engaged)相比,与该环相连的β折叠(βD2)发生了平移与旋转,这一运动被认为是R134重新定向所必需。该环构象也可被scFv16稳定,从而为其能够稳定无核苷酸复合物提供一种结构解释。
在核苷酸结合口袋内部,AHD开启促使αF向Gβ方向回撤,并导致α1的动态性增加,这一趋势也可在分子动力学(MD)模拟中得到进一步支持。与unlatched态相比,primed态的AHD构象波动显著降低,提示Gβ1对AHD提供了更稳定的协调约束。与“primed态对GDP亲和力最低”的推断一致,MD轨迹显示该状态下GDP表现出最强的构象异质性:在4条轨迹中,鸟嘌呤基团被逐出,核苷酸仅通过磷酸基团相互作用维持结合;另有1条轨迹中核苷酸完全解离。
2.3.5 无核苷酸态MOR-Gi
向无核苷酸态推进的最后一项重排表现为P环发生塌陷并收缩约4Å,从而不利于GDP重新结合。与此同时,在G蛋白异源三聚体中还可观察到一个逐步变化的整体几何趋势:比较从受体游离状态到无核苷酸状态,Gαi与β亚基之间的相对夹角逐渐变得更“平坦”,总变化幅度约15°。这一结果与受体依赖的G蛋白激活“杠杆臂”假说相一致。
2.4 G蛋白激活过程中的配体动力学
对这些cryoEM结构进行比较后可见,在10张密度图中的8张里都能观察到纳洛酮与洛哌丁胺的清晰配体密度。阿片受体中拮抗剂与激动剂的一个重要区分特征,是配体在正构结合口袋内的结合“高度”不同:拮抗剂通常呈现相对更“浅”的结合姿态,而激动剂更“深”。与这一规律一致,在两种“非活化”的MOR构象中,纳洛酮在受体内呈现浅结合姿态,与后续各状态形成对照。相反,洛哌丁胺在正构口袋中的位置并未表现出明显移动,这可能与高分辨率的洛哌丁胺结合中间态均对应“类活化”受体构象有关。总体而言,尽管配体姿态变化较为细微,仍可观察到配体结合方式与GDP协调程度之间的相关性,提示配体构象与核苷酸结合状态在激活过程中存在耦联关系。
此外,此前用于解析GDP结合复合物的主要策略是先将复合物去除核苷酸,再补加GDP,即“再结合GDP”(re-bound GDP)方案。为排除由该流程可能引入的结构伪影,研究还采用“恒定GDP”(constant GDP)条件进行验证,即纯化纳洛酮结合或洛哌丁胺结合的MOR复合物时,使用GDP结合的Gαi异源三聚体并在全部纯化缓冲液中维持200-500μM的GDP。该条件下复合物能够顺利经受纯化流程,并得到与上述各状态一致的三维重构结果。与提出的机理预期相符,纳洛酮结合的latent态以及洛哌丁胺结合的“开放AHD”构象获得了最高分辨率的重构。总体来看,这些结果有力支持了原始实验流程的可靠性以及所提出的结构与机理解释。
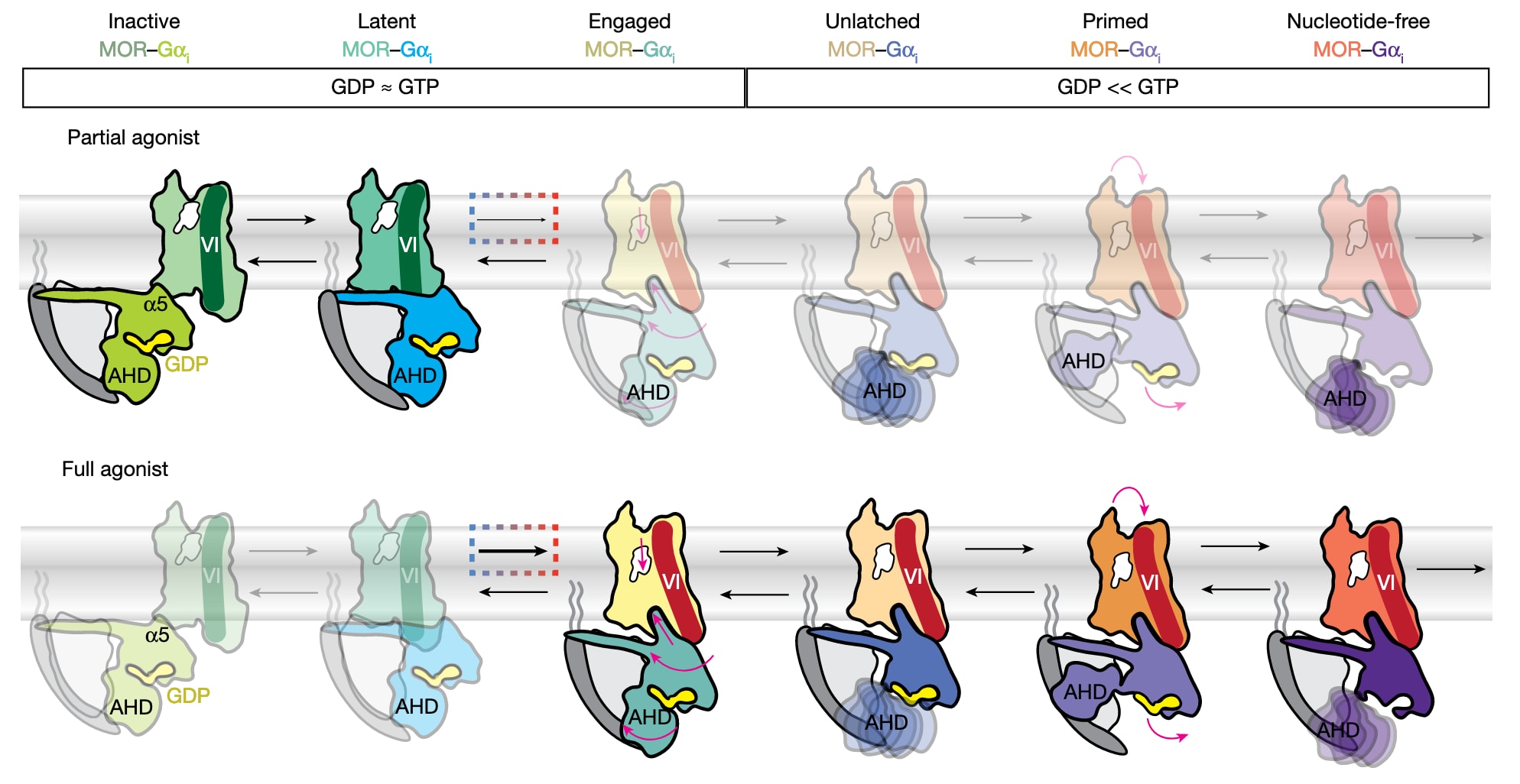
图4 | 展示了|MOR介导的G蛋白激活模型:配体效能通过结合姿态与构象稳定性调控GDP释放速率。 该模型提出,配体效能决定其在正构口袋内的结合姿态,并选择性稳定特定构象状态,从而调控GDP释放速率。数据支持的关键观点是,效能主要由复合物从信号沉默的latent态向engaged态的转变所驱动:这一转变产生典型“类活化”的MOR构象,并伴随α5螺旋延伸,尽管此时AHD仍保持闭合。engaged态在结构上已为后续AHD开启做好准备,而AHD开启对GDP释放至关重要。latent态的稳定可能降低核苷酸交换速率,使体系更接近受体游离的GDP-Gαi1β1γ2所表现的基础活性;相反,沿下游状态推进可加速GDP释放,从而增强G蛋白激活。
3 总结
相关研究提供了MOR-G蛋白激活通路早期阶段的一组构象快照,从而为理解MOR药理学的基础机制提供了结构线索。结构比较显示,在GDP释放之前,配体结合姿态与G蛋白构象状态之间存在直接相关性:在“非活化”受体构象中,纳洛酮在正构结合口袋内呈现浅结合姿态;而在“活化”构象中,纳洛酮与洛哌丁胺均在口袋内结合得更深。这一现象提示,效能更高的配体更倾向于稳定有利于核苷酸交换的受体构象。
在通路起始阶段,研究解析了latent态MOR-G蛋白复合物结构,捕捉到一种早期相互作用状态。与既往结构不同,该状态下MOR通过全部三个细胞内环(ICL1、ICL2、ICL3)与G蛋白相互作用。此前核磁共振研究提示,激动剂结合会增强ICL1的动态性,而TM6的完全重排则需要G蛋白存在,这与latent态所揭示的“早期结合-后续重排”的分步特征相呼应。随后,研究解析了GDP结合的三元复合物engaged态结构,其特征包括α5螺旋完全延伸且AHD保持闭合。尽管如此,分子动力学(MD)模拟显示,在多数轨迹中AHD可出现短暂开启,提示α5延伸、AHD开启与GDP释放更可能是依次发生,而非同步完成。
在更靠近GDP释放的阶段,研究还捕捉到两个AHD开启的状态,即unlatched态与primed态,代表GDP释放前的直接步骤。针对unlatched态的MD结果表明,AHD高度动态,其与RHD的随机碰撞可能导致核苷酸通道被间歇性遮挡,从而限制GDP释放。primed态则指向一种类似“活门”的机制:当AHD回撤时,αF螺旋同步回撤,使得GDP的协调达到各状态中最动态的程度;在某条primed态MD轨迹中,这一过程最终导致GDP被逐出。此外,某条unlatched态MD轨迹会逐渐过渡到primed构象,同样引发GDP释放。由于在所有含GDP的样品中都观察到了“AHD开启”构象,该状态可能比以往认识更为常见。四种GDP结合中间态的存在也提示,在实验条件下GDP结合具有可逆性,并不必然导致三元复合物解离。
综合这些结果,研究提出一个模型:配体效能与跨越G蛋白激活限速步骤(GDP释放)的能力相关,其本质是通过改变受体构象平衡来调控核苷酸交换。拮抗剂与部分激动剂会使MOR-Gi复合物停滞在latent态,该状态中多数GDP协调相互作用仍保持完整,因而可能对应较低的GDP释放速率。相较之下,激动剂洛哌丁胺更容易推动复合物进入engaged态,该状态在结构上已为AHD开启及随后的GDP释放做好准备,并降低GDP再结合的可能性。因此,由洛哌丁胺稳定的复合物平均表现出更低的GDP亲和力,而由纳洛酮稳定的复合物则相对更高。值得注意的是,即使在“恒定GDP”(constant GDP)的纯化流程中,仍能观察到engaged、unlatched与primed等“AHD开启”状态,提示这些状态确可再结合GDP,并且在实验条件下依然稳定。总体而言,GDP释放可被配体通过调控从非活化到latent、再到engaged、unlatched与primed等一系列状态所构成的构象景观而控制。进一步外推,反向激动剂可能主要通过稳定latent态而发挥作用,这或可解释为何与激动剂不同,MOR拮抗剂的结合不受核苷酸存在与否的影响,并与近期在KOR体系中的观察相一致。
在构象通路的组织方式上,研究更倾向于将G蛋白激活所需的构象重排视为线性序列,但也不排除这些状态之间还可通过更多分支路径相互连通。尽管无法完全排除配体在无核苷酸状态之后仍可能产生影响,实验并未观察到GTP结合亲和力的提升;相反,超激动剂洛哌丁胺对应的GTP亲和力还出现下降,进一步提示激动剂的主要作用机制是促进核苷酸释放而非增强GTP结合。
这些发现与单分子Förster共振能量转移研究形成互补,后者指出配体效能与GDP结合的“预激活”构象与完全“活化”构象之间平衡的改变相关。同时,这些结构也与近期报道的GTP结合后MOR-Gαi复合物构象高度相似:GDP结合的latent态接近于“G-ACT-2/3”,engaged态接近“G-ACT-1”,primed态则与“primed”构象相符,并呈现出相近的整体界面重排以及AHD逐步闭合的趋势。主要差异在于,当前结构仅在SwII中观察到细微变化,且未见亚基解离;不过,GDP释放在构象变化与状态序列上与GTP结合高度相似并不令人意外。总体而言,该研究通过揭示配体能够稳定不同构象比例,从而造成不同程度的核苷酸交换,为理解MOR配体效能提供了结构基础,并强调了高分辨率的核苷酸结合GPCR-G蛋白三元复合物结构在解析中间态方面的独特价值,其结构细节层面的信息密度难以由其他方法达到。鉴于功能选择性的中间态可导向特定药理学结局,这一构象调控模型也可能在GPCR超家族中具有更广泛的适用性。