NC 2025 | 基于结构的宏环肽设计用于生成针对G蛋白偶联受体的功能性抗体
今天介绍的是发表在 Nature Communications 上的一项研究,该工作围绕长期以来难以攻克的GPCR抗体开发问题,提出了一种具有普适潜力的新策略。研究通过理性设计宏环肽,在结构层面模拟GPCR胞外环的三维构象,并利用与哺乳动物系统发育距离较远的鸡作为免疫宿主,成功获得了高特异性、高一致性的抗GPCR抗体。作者以在多种癌症中高表达的神经降压素1型受体NTS1为模型靶点,系统展示了从计算建模、肽段设计到抗体功能验证的完整流程。研究不仅证明了宏环肽连接臂设计对抗体识别构象和功能活性的决定性作用,还获得了一种能够拮抗受体信号通路的功能性抗体。该工作为突破GPCR抗体免疫原性不足和构象依赖性强的瓶颈提供了清晰范式,对诊断工具开发和抗体药物设计具有重要启发意义。

获取详情及资源:
0 摘要
G蛋白偶联受体(GPCRs)在多种病理生理过程中发挥关键作用,因此成为药物开发及相关诊断工具研究中极具吸引力的靶点。尽管小分子药物已成功用于靶向GPCRs,但稳定可靠的抗GPCR抗体的制备仍然面临重大挑战。为解决这一问题,研究提出了一种新策略,利用宏环肽作为免疫原,模拟GPCR胞外环的三维结构,并选择在遗传上与哺乳动物差异较大的鸡作为免疫宿主,以产生抗原特异性抗体。研究以亲和力较高的神经降压素1型受体(NTS1)为靶点,该受体在多种人类癌症中高表达,且与不良预后密切相关。基于已报道的NTS1晶体结构,通过分子建模和预测分析软件对宏环表位模拟物及连接臂的选择进行理性设计。研究结果特别强调了连接臂在肽宏环化过程中的关键作用,其决定了所获得抗体是否能够发挥拮抗活性。总体而言,该策略为构建具有空间特异性的高效抗GPCR抗体提供了重要手段,并在诊断和治疗应用方面展现出良好的潜力。
1 引言
G蛋白偶联受体(GPCRs)是膜受体中规模最大的一类,在细胞生理功能和稳态调控中发挥核心作用。GPCRs在与内源性配体结合后,可激活多种信号通路,并通过G蛋白及其他细胞内效应分子引发多样化的细胞反应。GPCRs的功能失调及其表达水平的变化与多种病理状态密切相关,尤其与肿瘤生长和癌症发生有关,因此被视为极具潜力的治疗靶点。目前已获批准上市的药物中,超过35%是以GPCRs为作用靶点。鉴于GPCR家族在药物研发中的重要地位,开发可靠的抗GPCR抗体不仅有助于受体的基础研究与功能表征,也有望作为具有广泛应用前景的治疗性分子,在肿瘤、炎症、代谢性疾病及感染性疾病等领域发挥作用。同时,GPCRs作为诊断、预后及预测性生物标志物的重要性也日益凸显,因此亟需建立稳健的方法来制备具有高特异性、高选择性和良好重复性的抗GPCR抗体。然而,选择性靶向GPCR的抗体制备始终面临巨大挑战。迄今为止,仅有三种抗GPCR抗体获得美国食品药品监督管理局批准,分别是靶向CCR4、CGRPR和GPCR5D的mogamulizumab、erenumab以及近期报道的talquetamab。抗GPCR抗体的制备依赖于合适抗原的选择及其呈递方式以诱导免疫反应,其中最常见的方法是利用适当抗原对动物进行免疫,通常选择哺乳动物作为宿主。然而,在涉及GPCR这类嵌入细胞膜的复杂膜蛋白时,抗原形式的选择成为最大难题。目前大多数抗GPCR抗体是以受体的N端结构域或胞外环(ECL1-3)作为免疫原获得的,但关键挑战在于识别并构建能够模拟天然构象的正确折叠表位。不同物种间GPCR序列和结构的高度同源性也降低了免疫原性,尤其当抗原来源物种与免疫动物亲缘关系较近时更为明显。此外,GPCR全长蛋白的表达通常较弱且持续时间短,同时由于胞外表面暴露受限,蛋白易发生错误折叠并导致表位可及性不足。为克服上述问题,该研究提出了一种新策略,利用宏环肽作为免疫原,模拟GPCR胞外环的三维结构,并在鸡体内进行免疫。由于鸡在遗传上与哺乳动物存在较大差异,该方法有望针对哺乳动物来源的表位诱导更强的免疫反应,从而获得稳定可靠的抗GPCR抗体。在这一概念验证研究中,研究选择高亲和力的神经降压素1型受体(NTS1)作为靶点。NTS1在多种癌细胞中高表达,并与不良预后密切相关。其天然配体神经降压素(NT)可由肿瘤细胞以自分泌方式释放,与NTS1结合后作为生长因子促进肿瘤进展、细胞侵袭和转移。此前已解析NTS1在激动剂结合状态下的晶体结构,为表位三维结构的预测提供了重要依据。研究系统描述了一种能够同时识别GPCR胞外环序列及其三维构象的抗体制备策略,并特别强调了用于免疫的宏环肽中连接臂在宏环化过程中的关键作用。所设计的肽段被预测可采用与NT结合状态下NTS1第二胞外环(ECL2)相对应的构象,由此获得的抗体能够选择性识别受体并表现出显著的拮抗活性,其功能性通过阻断NT诱导的下游信号通路得到验证。而对连接臂进行轻微修饰后,所得抗体虽仍具有良好的特异性和选择性检测能力,适用于诊断应用,但不再具备拮抗活性。总体而言,该研究提出的技术为靶向GPCR抗体的开发提供了一种重要进展。
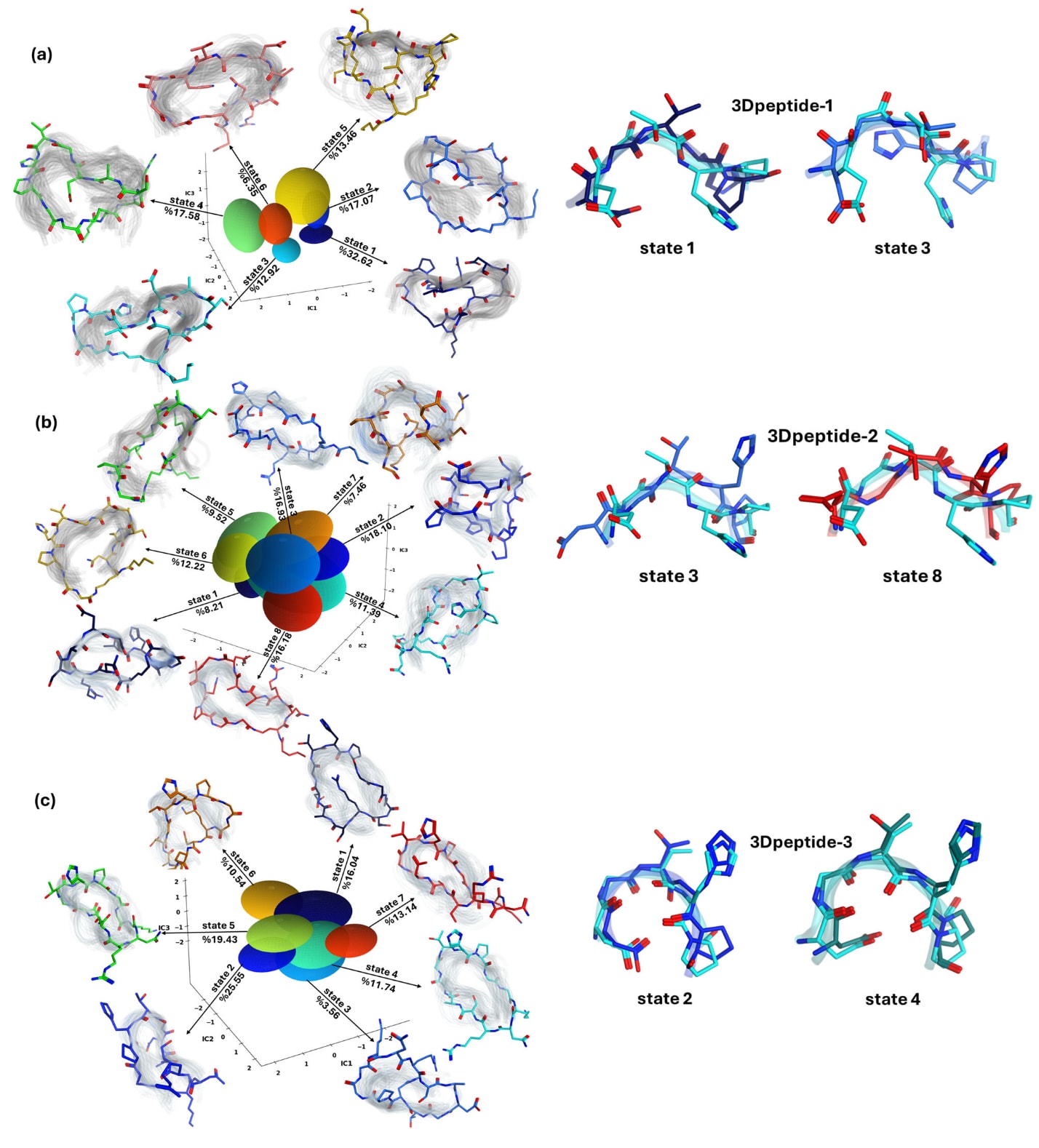
图1 | 3D肽的预测亚稳态构象及其与ECL2主链的叠合结果。 行(a–c)分别对应3Dpeptide-1、3Dpeptide-2和3Dpeptide-3。每一行的中央面板展示了分子动力学数据在三维tICA空间中的嵌入结果,由高斯隐马尔可夫模型识别的亚稳态构象以彩色椭球表示,并标注其所占比例。围绕该嵌入图的是每个状态中随机选取的50个构象的带状结构表示,并叠加于整个构象集合的半透明密度分布之上。每一行最右侧的面板显示了与激动剂结合状态下rNTS1受体胞外环2(ECL2)主链进行叠合的两种最低RMSD质心结构,以棒状形式表示,其颜色与对应椭球的颜色一致,rNTS1的ECL2主链以青色带状结构表示。
2 结果
2.1 NTS1第二胞外环肽模拟物的理性设计与合成
为开展该研究,针对NTS1的每一个胞外环(ECL1,ECL2和ECL3),宏环抗原的设计主要基于两个标准:其一是环的尺寸,即参与成环的氨基酸数量及用于实现宏环化的连接臂。为覆盖尽可能广泛的化学空间并获得构象多样性,研究共评估了123种连接臂,并将其分为7类,包括芳香族、联苯、吲哚、哌嗪、线性结构、氨基酸类及其他类型。连接臂的筛选依据包括以下方面:结构柔性与化学骨架的多样性、合成的可行性或商业可得性、是否含有两个适合成环的官能团,以及是否含有可用于与KLH偶联的游离巯基或能够在连接臂邻近位置引入半胱氨酸。针对每一个胞外环,分别考虑9至12个氨基酸,从而在计算机中共评估了近1500种宏环结构。在采用多丙氨酸模型宏环进行能量最小化并结合人工目视检查后,最终选取了24种连接臂用于进一步的分子动力学研究,共生成288种宏环结构(ECL1、ECL2和ECL3,每种长度9至12个残基,24种连接臂×3个环×4种长度)。综合后续的生物学实验结果、分子建模分析以及下文所述的筛选标准,研究最终将重点放在ECL2上。
该研究选用的抗原决定簇肽序列由11个氨基酸组成(N-R-S-A-D-G-T-H-P-G-G),该长度处于表位的理想范围内,通常认为10至20个氨基酸最适合诱导免疫反应。结构明确的表位模拟物的选择与理性设计综合考虑了多个因素,包括胞外结构域的生物学相关性、其在受体功能中的作用(例如ECL2已被证实参与受体活性)、肽段的免疫原性、激动剂结合状态下大鼠NTS1受体晶体结构的可获得性,以及肽合成的可行性。随后,利用Molecular Operating Environment(MOE)软件中集成的多种算法进行计算设计与模拟,以筛选适合肽环化的连接臂,并预测所合成宏环肽的柔性和亲水性。
肽宏环的设计流程首先通过构象搜索确定初始结构状态,随后进行分子动力学模拟以探索体系的动态行为。接着,采用时间滞后独立成分分析(tICA)识别慢速动力学过程,并进一步通过高斯隐马尔可夫模型(GHMM)优化,对亚稳态构象进行聚类并计算其转移过程。整个流程最终通过全面验证和系统分析得以完成,从而深入揭示体系的构象行为与动力学特征。基于该流程,研究鉴定并合成了三种构象受限的肽,通过三种不同的连接方式闭环,以模拟ECL2的三维结构,分别为侧链S-赖氨酸闭环的3Dpeptide-1、侧链S-鸟氨酸闭环的3Dpeptide-2以及主链S-赖氨酸闭环的3Dpeptide-3。同时,还合成了一条线性肽(Lin-peptide)作为对照。
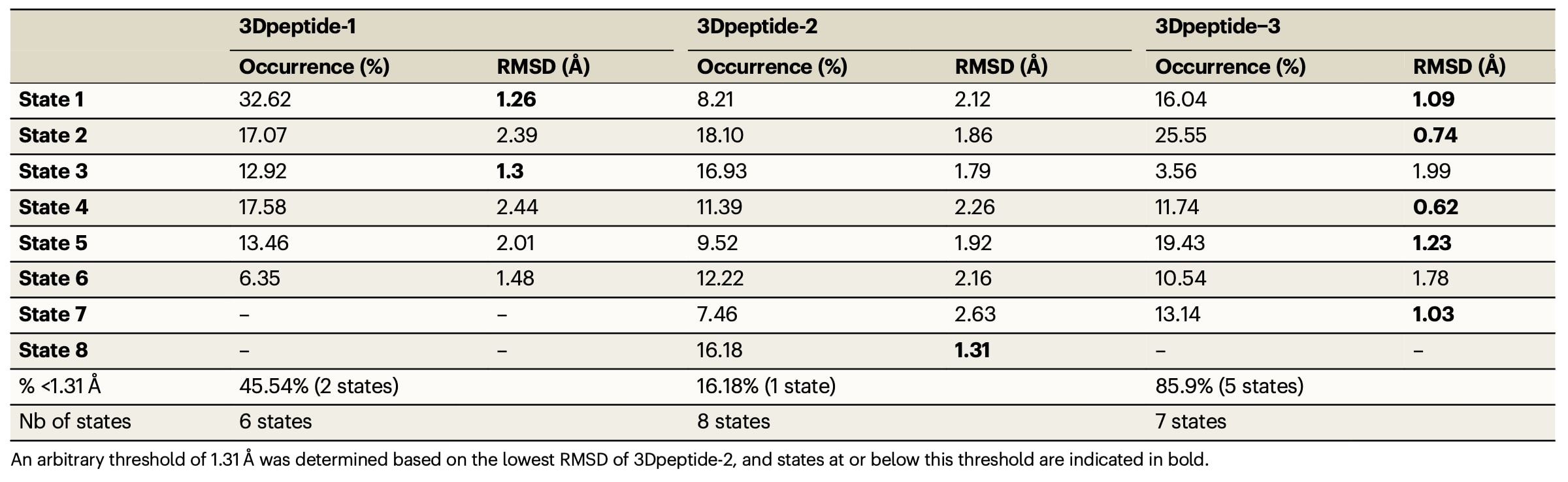
针对所有三种3D肽均开展了分子建模分析,以预测哪一种连接臂能够更好地再现ECL2的构象特征。所有预测得到的构象状态及其对应的出现比例以彩色球体形式展示于图1中。为评估空间肽对ECL2的模拟程度,研究计算了每一种3D肽在各预测状态下与rNTS1晶体结构中ECL2之间的原子位置均方根偏差(RMSD)。RMSD的计算基于环中暴露程度最高的5个氨基酸(D-G-T-H-P)的α碳和β碳,并在MOE软件序列编辑模块的Align/Superimpose界面中完成叠合。RMSD值最低的构象被认为是最接近天然ECL2的模拟物。为便于比较,研究设定1.31 Å作为阈值,该值对应三种肽最低RMSD中的最大值(3Dpeptide-1为1.26 Å,3Dpeptide-2为1.31 Å,3Dpeptide-3为0.62 Å)。
隐马尔可夫模型分析显示,3Dpeptide-1存在6种亚稳态构象,其中2种状态的RMSD小于等于1.31 Å,合计占据45.54%的概率。其中状态1不仅具有最低的RMSD值(1.26 Å),同时也是3Dpeptide-1中稳定性最高的构象。对于3Dpeptide-2,共预测得到8种构象状态,仅有1种状态满足阈值要求,其出现概率为16.18%。相比之下,3Dpeptide-3共有7种预测构象,其中5种状态的RMSD低于阈值,累计占比达到85.9%,因此被预测为最有可能采用与激动剂结合状态下rNTS1受体ECL2相似的三维结构。进一步分析表明,3Dpeptide-3中RMSD最低的两种状态(0.62 Å和0.74 Å)在主链构象上与rNTS1的ECL2表现出最佳叠合效果。图1右侧面板展示了每一种肽中RMSD最低的两种预测构象与激动剂结合状态下rNTS1的ECL2主链的叠合结果。
值得注意的是,这些构象占据比例的排序与实验检测结果高度一致,表明在ECL2模拟构象中停留时间越长,其与受体的结合能力越强。隐含时间尺度曲线在约6 ns处达到平台,CK检验显示预测与观测到的转移概率高度重合,验证了动力学模型已充分收敛并满足马尔可夫性。这些结果为实验中观察到的效力排序提供了统一而清晰的机制性解释。
表1 | 各肽构象状态的出现比例及其对应的RMSD计算值。
2.2 抗NTS1抗体的亲和力与选择性
在分别使用四种肽(Lin-peptide、3Dpeptide-1、3Dpeptide-2和3Dpeptide-3)进行免疫后,从鸡体内提取并纯化获得多克隆抗体(IgY),最终制备出四种不同的抗NTS1抗体,分别命名为IgY-Lin、IgY-3D-1、IgY-3D-2和IgY-3D-3。IgY-Lin主要被设计用于识别线性表位(N-R-S-A-D-G-T-H-P-G-G),而其余三种抗体则旨在优先识别该序列所呈现的三维构象。为验证这些抗体与相应肽段的结合能力,首先通过酶联免疫吸附实验在固定肽条件下对所有抗体进行检测。通过将抗体与逐渐增加浓度的对应游离肽(0至10 μM)预孵育,生成抑制曲线,从而以剂量依赖方式阻断抗体对固定肽的结合。结果显示,四种抗体抑制50%结合所需的浓度相近,IgY-Lin的IC50为2.2 ± 0.6 nM,IgY-3D-3为7.8 ± 2.3 nM,其余抗体介于两者之间。上述结果表明,各抗体对其对应肽段具有相似的检测能力,同时验证了免疫策略的有效性。
随后,为分析这些抗NTS1抗体的选择性,研究进一步开展了竞争性ELISA实验,以四种设计肽作为固定诱饵,并加入固定浓度为10 μM的各类肽以竞争IgY的结合。结果显示,所有抗体均可被其对应肽有效阻断,阻断效率相近,分别为IgY-Lin 95% ± 3.5、IgY-3D-1 100% ± 0.1、IgY-3D-2 99% ± 1.2以及IgY-3D-3 97% ± 2.3。IgY-Lin与其线性肽的结合在加入三种3D肽后可被部分抑制,其中3Dpeptide-1的抑制效果最为明显,阻断率可达62%,提示其存在一定程度的交叉反应性。相比之下,IgY-3D-1仅能被其对应的3Dpeptide-1有效阻断,而不受其他三种肽的影响,显示出高度特异性。IgY-3D-2和IgY-3D-3的特异性则相对较低,这两种抗体与固定肽的结合可被其他肽部分置换,包括线性肽。综合来看,与线性表位相比,针对3D肽获得的抗体在识别三维表位方面表现出更显著的特异性。
研究还进一步评估了肽序列本身在抗体识别中的重要性。通过将预测为主要暴露于表面的氨基酸残基(D-G-T-H-P)逐一替换为丙氨酸(A),即进行丙氨酸扫描分析,系统考察各残基对抗体结合的贡献。该分析聚焦于最能模拟ECL2三维结构的两种宏环肽,即3Dpeptide-1和3Dpeptide-3。与野生型3Dpeptide-1的结合情况相比(IC50为3.2 ± 0.6 nM),IgY-3D-1对所有突变肽的亲和力均显著降低,其中D261A、G217A和T218A突变影响尤为明显,对应的IC50值分别升高至2447 ± 852 nM、1286 ± 354 nM和708 ± 194 nM。相较之下,IgY-3D-3对3Dpeptide-3中这些突变的敏感性较低,抗体结合能力未出现显著下降。与野生型3Dpeptide-3的IC50值7.8 ± 2.2 nM相比,T218A和H219A突变甚至略微增强了IgY-3D-3与肽段的结合能力,其IC50值分别为0.72 ± 0.04 nM和0.58 ± 0.07 nM。
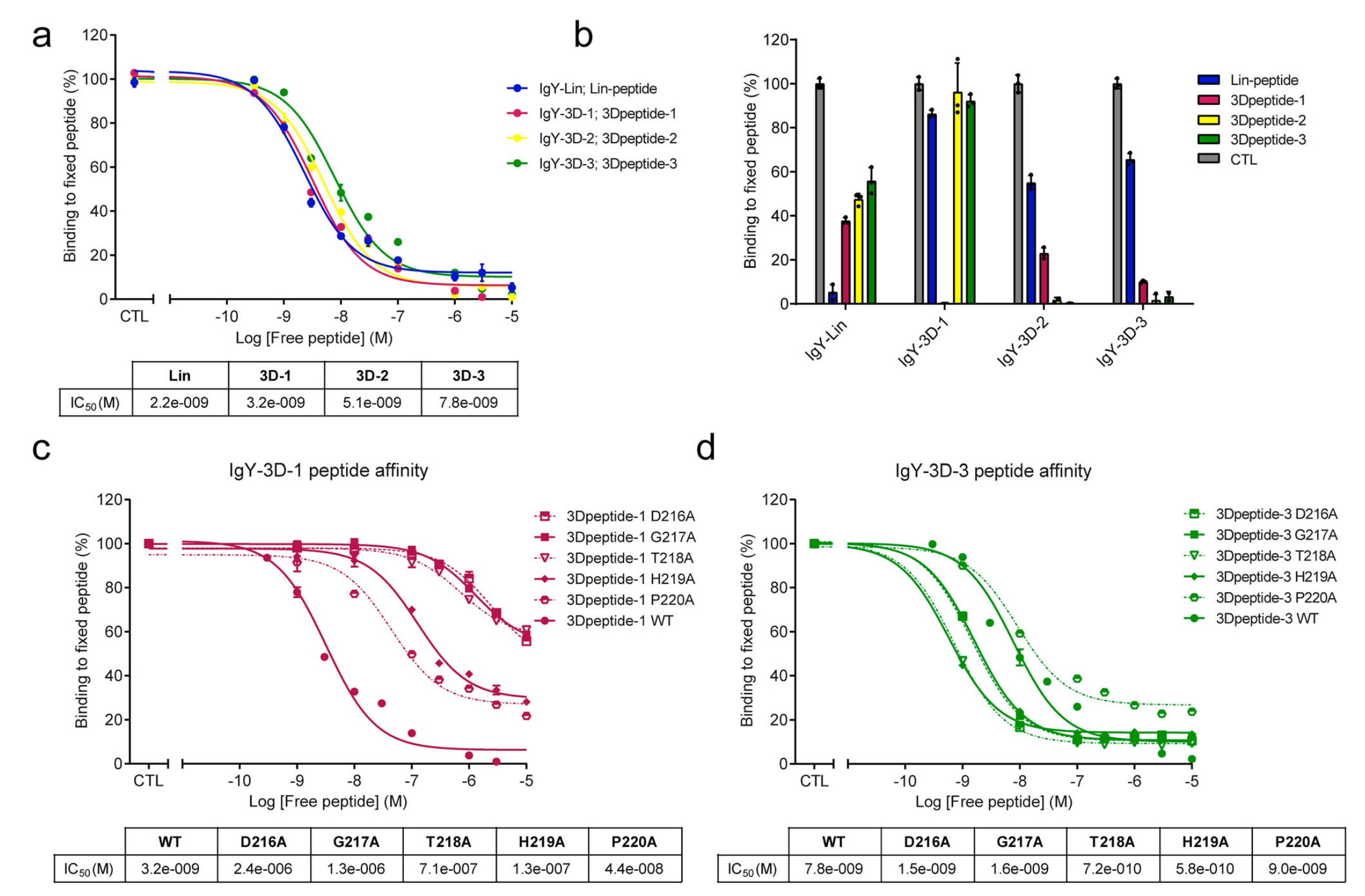
图2 | 抗NTS1抗体的亲和力与特异性。 a 图示通过基于ELISA的方法,在逐渐增加游离肽浓度的条件下,各IgY抗体(IgY-Lin、IgY-3D-1、IgY-3D-2和IgY-3D-3)与其对应固定肽结合的抑制曲线,吸光度在450 nm处测定。仅含抗体而不加入游离肽的对照样品被设定为100%检测信号,其余样品均以对应对照为基准进行百分比归一化。b 图示在固定浓度为10 μM的不同游离肽竞争条件下,各IgY与其对应固定肽结合的抑制情况。阳性对照(灰色柱)表示各IgY对其特异性固定肽的完全检测。所有在450 nm处获得的原始数据均相对于不含肽的对照进行归一化,该对照定义为100%检测信号。c、d 分别展示IgY-3D-1(c)和IgY-3D-3(d)对不同突变宏环肽的亲和力,通过在逐渐增加五种指定突变游离肽(D216A、G217A、T218A、H219A、P220A)及野生型(WT)3Dpeptide-1和3Dpeptide-3浓度的条件下生成抑制曲线。所有曲线均采用三参数非线性回归进行拟合,用于计算IC50值,相应拟合结果以表格形式列于图下。数据以平均值 ± 标准差表示,每个实验包含2–4个技术重复,且每张图至少代表两次独立的生物学重复(n = 2)。每一个孔对应一次独立的技术重复,单个实验中以单个孔作为统计分析的基本单位。原始数据已作为Source Data文件提供。
2.3 抗体验证:在细胞及脑组织中检测NTS1靶蛋白
随后评估了这些抗NTS1抗体在整体细胞环境中检测完整靶蛋白的能力(图3)。首先,在稳定表达rNTS1受体的HEK293A固定细胞中进行免疫荧光显微观察,该受体在其N端胞外结构域带有血凝素(HA)标签(图3a)。在相同条件下,以单一浓度5 μg/ml检测四种抗NTS1抗体(IgY-Lin、IgY-3D-1、IgY-3D-2和IgY-3D-3)。同时使用小鼠抗HA抗体进行共定位分析,以验证受体的亚细胞分布。信号分别通过与鸡IgY结合的ATTO488标记二抗和与小鼠抗体结合的Alexa Fluor 647标记二抗进行检测,两种荧光染料的激发与发射光谱彼此区分且无重叠。对于IgY-3D-1和IgY-3D-3,z轴叠加的共聚焦显微图像显示两种荧光信号高度重合(图3a)。在相同抗体浓度下,IgY-Lin仅产生中等强度信号,而IgY-3D-2几乎无法检测到NTS1。加入各自对应肽后,免疫检测信号可被完全抑制(图3a右侧面板)。
为进一步确认这些抗体识别目标蛋白的有效性,对所有重复实验进行了共定位分析。针对每一次重复,计算Pearson相关系数和重叠系数(488/647),并绘制散点图以展示rNTS1与HA信号之间的相关性(图3a中间面板)。散点图若主要向x轴聚集,表示以HA信号为主而缺乏rNTS1检测,这一现象在IgY-3D-2中尤为明显。相比之下,IgY-3D-3的散点图显示出良好的相关性,被视为明显的共定位。所有条件下Pearson系数和重叠系数的汇总结果分别见图3b和图3c。仅从Pearson系数分析来看,所有抗NTS1抗体产生的信号均与HA信号共定位,数值介于0.5至1.0之间,提示未出现明显的非特异性信号。然而,不同抗体的信号强度存在显著差异,重叠系数分析进一步表明IgY-3D-2无法正确检测rNTS1受体(图3c)。其平均重叠系数低于0.5(mean = 0.27 ± 0.08),反映缺乏共定位。相反,IgY-3D-1和IgY-3D-3产生了强烈的检测信号,几乎完全与HA信号重合,其重叠系数分别为0.98 ± 0.19和0.97 ± 0.11。IgY-Lin检测rNTS1的能力相对较弱,不同重复之间的重叠系数波动较大(0.89 ± 0.3)。
已有研究表明,使用醛类固定剂处理细胞或组织可能改变蛋白表位的构象,从而导致天然蛋白中可及的抗原决定簇在活细胞与固定细胞中暴露方式不同。为评估这些抗体在识别天然构象NTS1方面的能力,研究在固定前先将活细胞与不同抗NTS1抗体及抗HA抗体在相同浓度下孵育,随后进行免疫荧光观察(补充图5a)。结果显示,IgY-3D-3是唯一能够在活细胞中检测到天然构象NTS1的抗体。相应地,对活细胞进行流式细胞术分析表明,仅IgY-3D-3在与抗HA抗体共标记的细胞中实现了信号检测,其中89.02%的细胞分布在Q1右上象限(补充图5b、c)。
为进一步验证这些抗NTS1抗体对内源性rNTS1受体的检测能力,研究在成年大鼠脑固定切片上进行了免疫组织化学分析。选择此前报道rNTS1高表达的两个脑区,即黑质和大脑皮层(图3d)。在黑质致密部,IgY-3D-1和IgY-3D-3可清晰检测到NTS1阳性的神经元胞体,呈现强烈染色(箭头所示)。相比之下,IgY-Lin虽能检测到NTS1,但染色较为弥散,难以清晰分辨单个免疫反应阳性的细胞。在听觉皮层中,同样可在Ⅱ-Ⅲ层及Ⅴ层观察到NTS1免疫反应阳性的神经元胞体。其中在Ⅴ层,NTS1表达细胞主要为锥体细胞,其胞体及顶树突均显示出明显的免疫标记信号(箭头所示)。
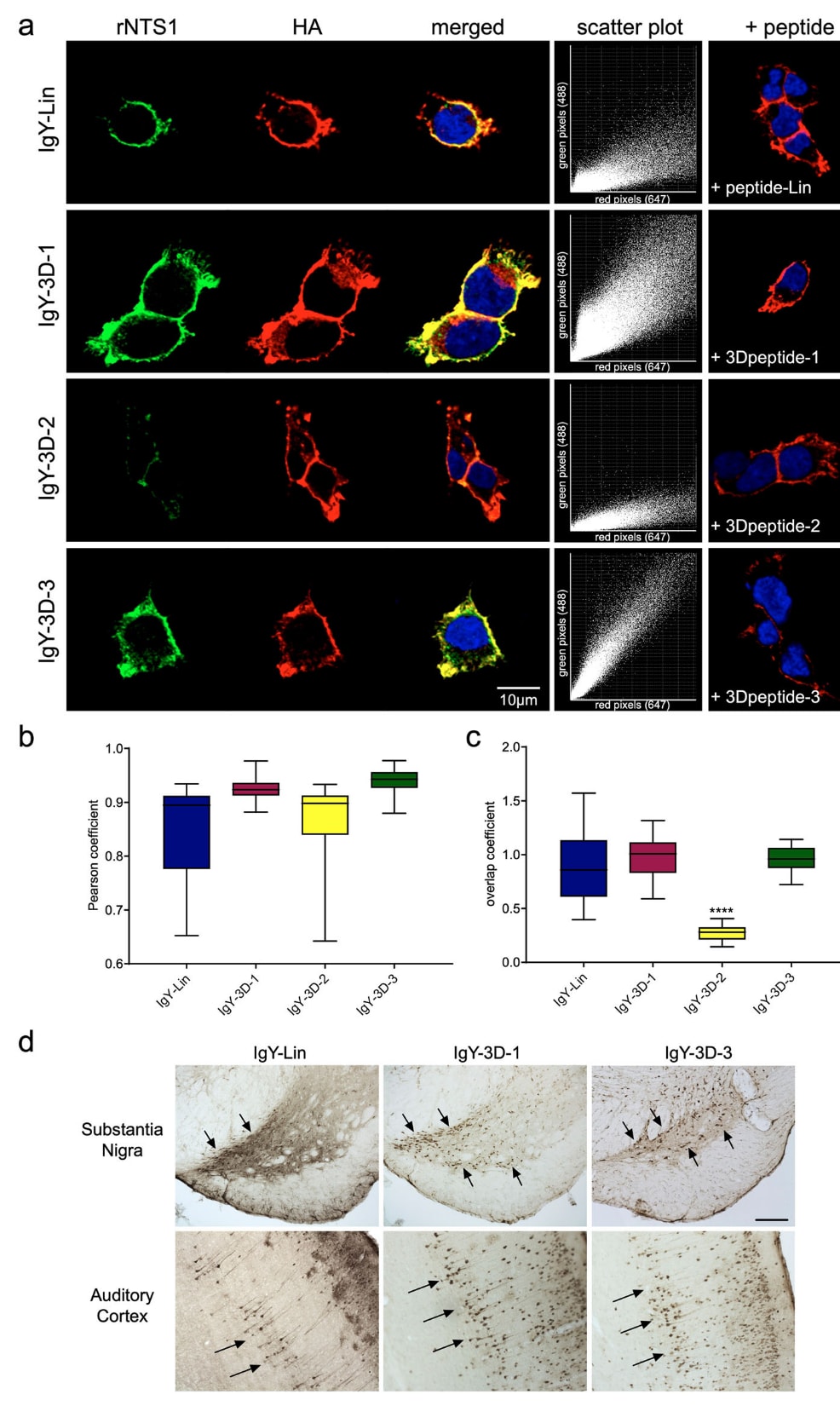
图3 | 通过免疫荧光显微镜和免疫组织化学对抗体进行验证。 a 固定的HEK293A细胞免疫荧光图像,该细胞稳定表达HA-rNTS1。绿色信号表示抗NTS1抗体(IgY-Lin、IgY-3D-1、IgY-3D-2和IgY-3D-3)对大鼠NTS1受体的检测,红色信号表示HA标签的特异性检测。右侧面板为阴性对照,通过将抗体与其对应肽按1:10比例预孵育后获得。Z轴堆叠图像使用共聚焦显微镜在60×物镜下采集,并通过3×数字放大以更清晰地显示感兴趣区域。b、c 分别为(a)中图像的Pearson相关系数和488/647重叠系数(绿色像素数/红色像素数)的计算结果。针对每一种抗体均给出了散点图,其中纵轴和横轴分别对应绿色(rNTS1)和红色(HA)信号。共定位分析基于从五个随机选取视野中获得的十个Z轴切片作为重复,并对Pearson系数(b)和重叠系数(c)进行定量。数据以最小值至最大值的箱线图形式展示,包含至少50个技术重复,代表两次独立的生物学实验(n = 2)。箱线图中,中心线表示中位数,箱体表示25%–75%分位区间,须线表示最小值与最大值。每一个视野中的单个Z轴切片被视为一次技术重复,在单个实验中以一个视野作为研究单位。统计分析采用单因素方差分析并结合Tukey多重比较检验,****表示P值<0.0001。d 成年大鼠脑固定组织切片中NTS1免疫反应的分布情况,使用抗NTS1抗体(IgY-Lin、IgY-3D-1和IgY-3D-3)进行检测。在黑质致密部,可见NTS1免疫反应阳性的神经元胞体和树突广泛分布(箭头所示),比例尺为200 μm。在大脑皮层中,第Ⅴ层的NTS1表达细胞主要为锥体细胞,其胞体和顶树突显示出强烈的免疫标记信号(箭头所示)。此外,在听觉皮层的Ⅱ层和Ⅲ层亦可观察到标记的神经元胞体。比例尺为150 μm。原始数据已作为Source Data文件提供。
2.4 抗体验证:通过生化分析检测NTS1
研究进一步通过免疫印迹(Western blot,WB)以及免疫共沉淀(IP)结合质谱(MS)的方法对这些抗NTS1抗体进行了系统表征(图4)。WB通常用于评估抗体是否能够识别变性状态下的抗原,这是由于裂解缓冲液中含有SDS,而IP实验则对蛋白构象高度敏感,需要抗体能够在天然状态下识别靶蛋白。上述实验均在来源于HEK293A细胞的全细胞裂解液中进行,这些细胞分别表达或不表达HA标签的rNTS1受体。
首先,使用抗HA抗体进行WB分析,在约45 kDa和55 kDa处检测到两条条带,对应此前报道的NTS1两种不同糖基化形式的预期分子量(图4a,红色虚线框)。采用IgY-3D-1和IgY-3D-3抗体同样检测到相似分子量的条带,其中IgY-3D-3在检测NTS1蛋白方面的效果明显优于IgY-3D-1(图4a,红色虚线框)。相比之下,IgY-Lin产生了明显的非特异性信号,在不表达NTS1的HEK293A细胞中也检测到相似分子量的蛋白条带,而IgY-3D-2则未能在WB中检测到NTS1。
随后,利用上述HEK293A细胞系并将抗NTS1抗体偶联至磁珠上开展IP实验(图4b)。所有3D-IgY抗体均能够有效地富集受体,其拉下效率与HA对照IP相当。IgY-Lin同样可实现受体拉下,但效率明显低于3D肽来源的抗体(图4b)。
为进一步确认IP实验中拉下的蛋白确为NTS1受体,研究将IP与质谱分析相结合进行验证(图4c、d)。首先,使用偶联IgY-3D-1和IgY-3D-3的磁珠从表达HA-rNTS1的HEK293A细胞中分离rNTS1受体,随后采用抗HA-HRP抗体进行WB检测(图4c)。洗脱样品经冷冻干燥后分别送往两个独立的质谱研究平台进行分析。结果从Rattus norvegicus数据库中明确鉴定出一条由胰蛋白酶消化产生的17个氨基酸肽段,对应于NTS1的C端结构域。所鉴定的肽段序列及其质谱图如图4d所示,完整谱图见补充图6,离子信息列于补充表3。此外,在与相应肽段预孵育后,抗NTS1抗体与受体之间的相互作用被显著阻断(图4d内嵌图)。
上述结果有力证明了IgY-3D-1和IgY-3D-3抗体均能够与预期的NTS1靶蛋白发生特异性相互作用。
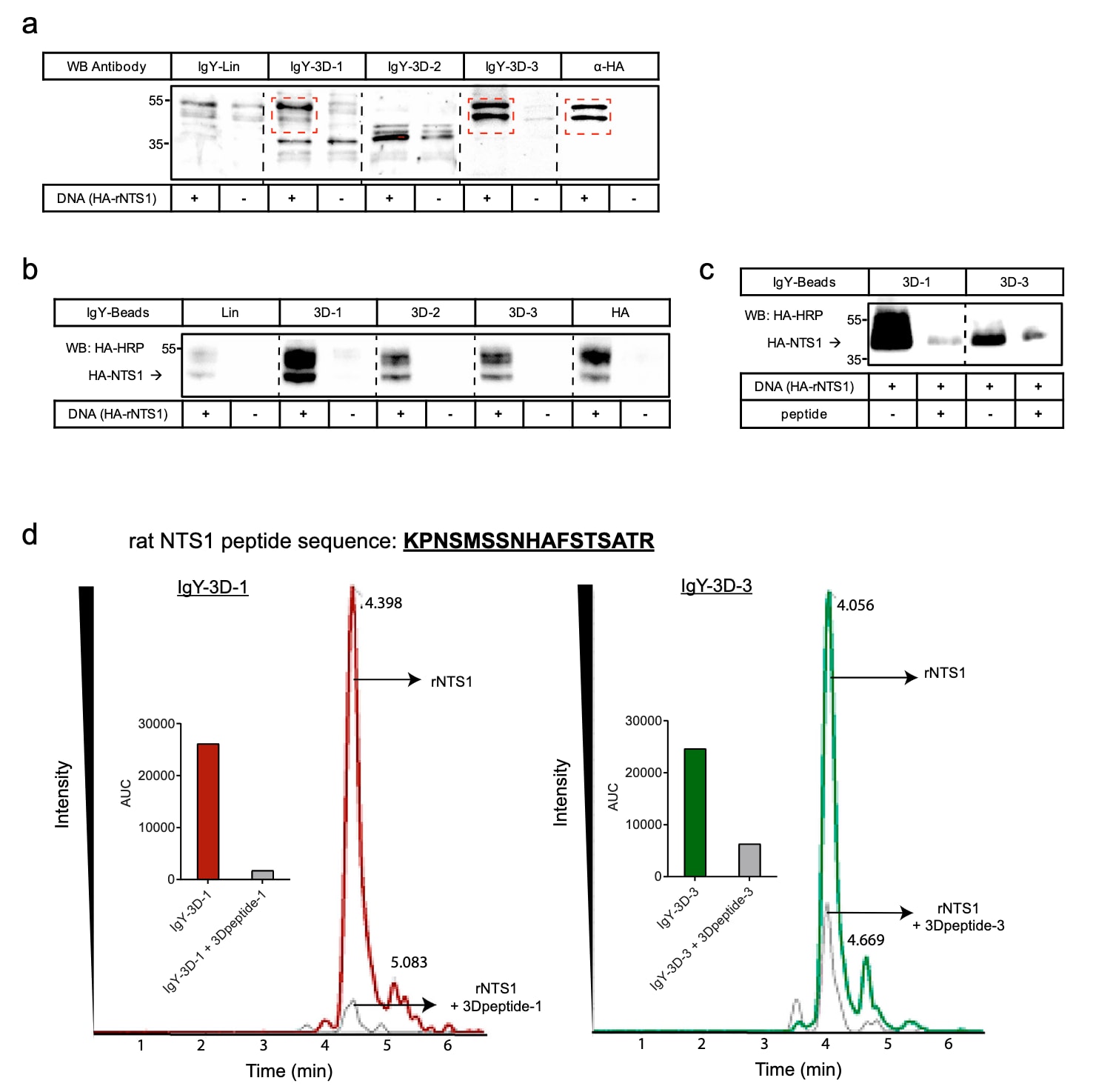
图4 | 通过WB、IP和MS对抗体进行验证。 a Western blot结果显示,在稳定表达HA-rNTS1或转染空载体作为对照的HEK293A细胞裂解液中,分别使用鸡源抗NTS1抗体(IgY-Lin、IgY-3D-1、IgY-3D-2和IgY-3D-3)及抗HA抗体检测大鼠NTS1受体。红色虚线框标示出对应rNTS1的信号条带。b 采用与磁珠偶联的各类鸡源抗体,对稳定表达HA-rNTS1的细胞进行NTS1的免疫共沉淀,并使用抗HA抗体进行免疫印迹检测。转染空载体的HEK293A细胞作为阴性对照。c 用于后续质谱鉴定的rNTS1免疫共沉淀样品的免疫印迹结果。阴性对照为在1:10比例下将IgY与其对应肽预孵育后进行的处理。d 采用IgY-3D-1(红色谱图)和IgY-3D-3(绿色谱图)进行质谱分析,得到的大鼠来源NTS1受体肽段的提取离子流色谱图(XIC)。与IgY对应肽预孵育的阴性对照以灰色曲线表示。(内嵌图)显示使用Peakview 2.2软件计算得到的各条件下曲线下面积(AUC)。所有数据均代表至少两次独立实验的结果。质谱分析在两个不同的研究机构中完成。原始数据已作为Source Data文件提供。
2.5 抗体的功能特性
尽管抗体被广泛用作研究膜蛋白的有力工具,但其同样具备调控蛋白功能的药理学潜力。基于这一点,研究进一步评估了这些抗NTS1抗体对NTS1受体活性的调节能力,这一特性在治疗应用中具有重要意义。由于NTS1与内源性激动剂神经降压素C端片段NT(8–13)结合的晶体结构显示,NT的配体结合口袋部分受到ECL2 β发夹结构的覆盖,研究首先检测这些靶向ECL2的IgY抗体是否能够竞争性地将放射性标记的NT从其结合位点置换下来。为此,将表达HA-rNTS1的HEK293A细胞膜与放射性标记的NT(125I-Tyr3-NT)共同孵育,同时加入逐渐增加浓度的IgY-Lin、IgY-3D-1、IgY-3D-2或IgY-3D-3(图5a)。结果显示,IgY-3D-3能够以剂量依赖方式有效置换125I-Tyr3-NT,其IC50为3.2 ± 0.7 μg/ml。相比之下,IgY-3D-1和IgY-Lin的置换能力明显较弱,IC50分别为36.5 ± 11.8 μg/ml和187 ± 94.9 μg/ml。IgY-3D-2几乎不具备竞争结合能力,这一结果与其无法在变性或天然状态下识别NTS1的特性一致。值得注意的是,按照该策略制备的IgY-3D-1和IgY-3D-3在不同批次、不同免疫鸡来源中均表现出稳定一致的识别特性,能够高效置换rNTS1结合的125I-Tyr3-NT,显示出良好的重复性。
在不同细胞类型中,NTS1受到NT刺激后可激活Gαs、Gαq、Gαi/o和Gα13信号通路,并招募β-arrestin 1和2。为判断这些抗NTS1抗体是否能够调节NTS1依赖的信号活性,研究首先采用基于细胞的均相时间分辨荧光(HTRF)方法,检测Gαs介导的环磷酸腺苷(cAMP)累积。具体而言,在稳定表达rNTS1的CHO-K1细胞中,以NT的EC50浓度进行刺激,并同时加入IgY-Lin、IgY-3D-1、IgY-3D-2或IgY-3D-3(图5b)。结果表明,仅IgY-3D-3能够显著抑制NTS1介导的cAMP生成。
随后,研究利用生物发光共振能量转移(BRET)生物传感器,在瞬时表达rNTS1的HEK293细胞中,系统评估这些抗NTS1抗体在结合受体后对Gαq、Gαi2、Gα13、GαoB、Gαz及β-arrestin 2信号通路的调节作用。结果显示,IgY-Lin、IgY-3D-1和IgY-3D-3均未表现出激动剂模式下的活性,既不能激活经典的Gαq和β-arrestin 2信号通路,也不能诱导NTS1与Gα13、GαoB、Gαi2或Gαz发生偶联。在拮抗剂模式下,IgY-3D-3则能够以浓度依赖方式有效阻断NTS1激活所涉及的全部信号通路(图5d、f及补充图7)。在BRET实验中,非肽类NTS1选择性拮抗剂SR48692被用作对照,其能够有效逆转NT在NTS1受体上的信号活性。在相同实验条件下,IgY-Lin和IgY-3D-1均无法抑制NT诱导的NTS1与G蛋白或β-arrestins之间的功能性偶联。
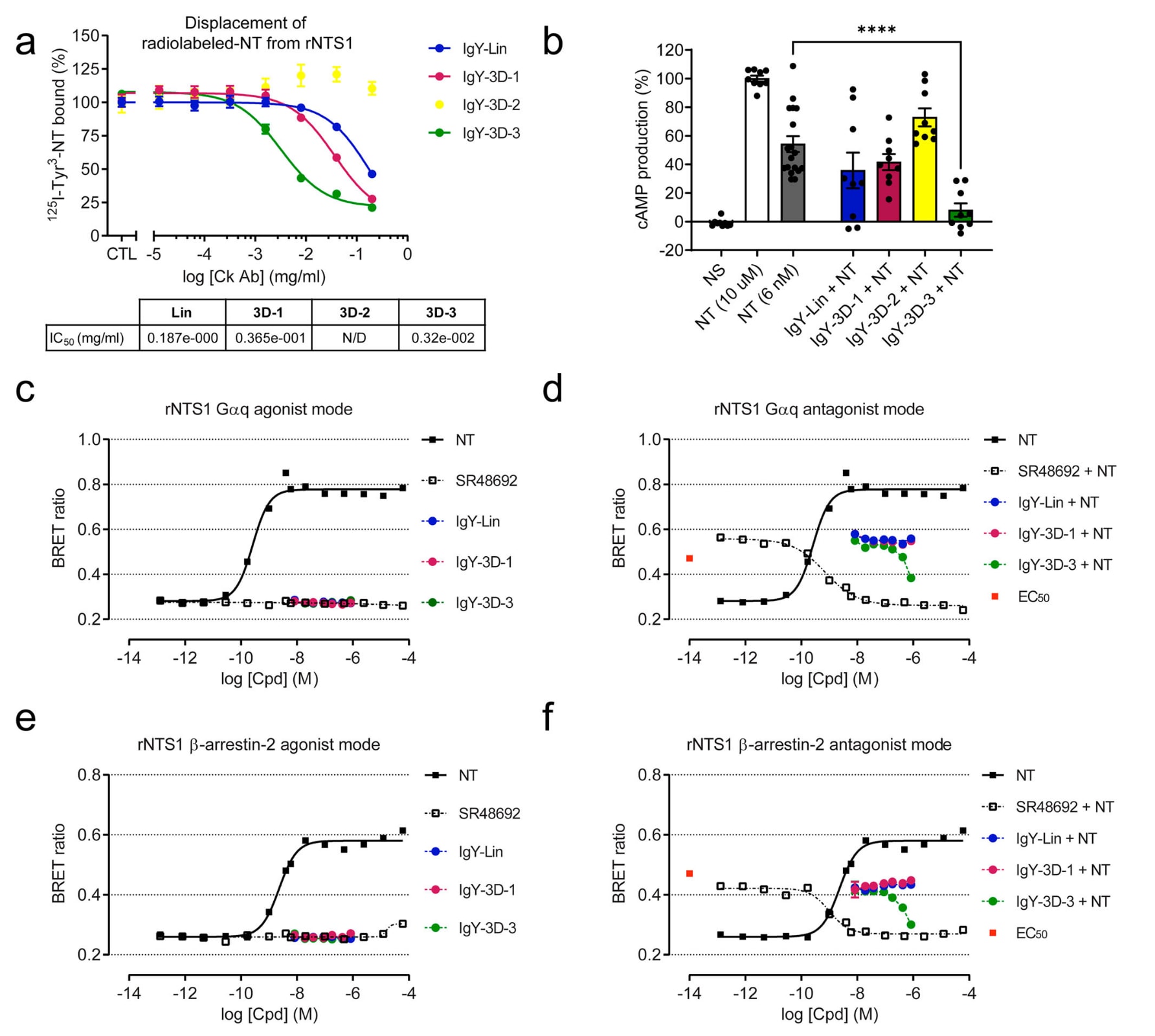
图5 | 抗体的功能特性。 a 鸡源抗体(IgY-Lin、IgY-3D-1、IgY-3D-2和IgY-3D-3)对放射性标记NT的置换曲线。曲线采用三参数非线性回归进行拟合,并由此计算图例中标注的IC50值。数据以平均值 ± 标准误表示,来源于两次独立生物学实验(n = 2,独立实验板),每个实验板中包含技术重复,每个孔视为一次技术重复。b IgY抗体对NT诱导的环磷酸腺苷(cAMP)生成的抑制作用。阴性对照为未刺激细胞(NS),所有抗体均在NT(EC50)存在的条件下进行检测。数据以平均值 ± 标准误表示,每个实验至少包含9个技术重复,代表两次独立的生物学实验(n = 2)。统计分析采用单因素方差分析并结合Tukey多重比较检验,****表示P值<0.0001(GraphPad在校正后的精确P值低于软件阈值时报告为<0.0001)。所示抗体在激动剂模式下对NTS1依赖的Gαq(c)和β-arrestin-2(e)信号通路的激活能力,以及IgY-Lin和IgY-3D-1/3在拮抗剂模式下对NT(EC50)介导的Gαq(d)和β-arrestin-2(f)信号通路的阻断能力。非肽类NTS1拮抗剂SR48692作为对照。所有曲线均采用四参数非线性回归分析。BRET实验以技术重复方式进行,结果代表两次独立的生物学实验(n = 2)。原始数据已作为Source Data文件提供。
2.6 抗体的交叉反应性与选择性
该研究充分利用了哺乳动物与鸡之间显著的系统发育差异,成功获得了两种稳健的抗NTS1抗体,即IgY-3D-1和IgY-3D-3,二者均针对rNTS1的ECL2区域设计。用于免疫鸡的ECL2宏环肽基于rNTS1的X射线晶体结构进行建模,其氨基酸序列在不同物种间高度保守,唯独在鸡(Gallus gallus)中存在差异(图6a)。尽管所设计肽与人源NTS1蛋白序列之间在苏氨酸和脯氨酸两个氨基酸位点存在差别,但将该设计肽与人源ECL2结构进行叠合后,其主链构象仍表现出高度相似性(图6b),提示大鼠与人源NTS1在三维构象上可能具有保守性。基于此,研究进一步评估了IgY-3D-1和IgY-3D-3在表达HA标记人源NTS1(hNTS1)的HEK293细胞中对hNTS1的检测能力。免疫荧光共聚焦显微观察(图6c)及基于全细胞裂解液的免疫共沉淀实验(图6d)显示,IgY-3D-1能够有效检测hNTS1,而IgY-3D-3则未表现出明显检测能力。相应地,IgY-3D-1在竞争置换结合NT方面同样表现出较强能力,其在hNTS1和rNTS1上的IC50值相近,分别为44.3 ± 20.6 μg/ml和36.5 ± 11.8 μg/ml(图6e)。值得注意的是,IgY-3D-3仍部分保留了置换hNTS1上结合NT的能力,其IC50为181 ± 91 μg/ml,但明显弱于其在rNTS1上的活性(3.2 ± 0.7 μg/ml)。
此外,研究还采用基于HTRF的细胞实验,在稳定表达hNTS1的CHO-K1细胞中检测Gαq依赖的肌醇一磷酸(IP1)累积。细胞在NT的EC50浓度下刺激,并同时加入IgY-3D-1或IgY-3D-3(图6f)。结果表明,两种抗体均可显著抑制IP1的产生,其中IgY-3D-1的抑制效果更为显著,这一趋势与放射性NT置换实验的结果一致。这些发现表明,IgY-3D-1在识别人源NTS1受体方面表现出更优的能力。
靶点选择性同样是治疗性和诊断性抗体开发中的关键因素。为此,研究进一步评估了IgY-3D-1和IgY-3D-3对与rNTS1高度相关的A类GPCR的识别能力,包括大鼠神经降压素2型受体(rNTS2)和大鼠apelin受体(rAPJ)。在过表达rNTS2、rAPJ或rNTS1的HEK293A细胞中分别开展WB分析(图6f)、天然条件下的IP实验(图6g)以及免疫荧光显微观察(补充图8a、b)。结果显示,IgY-3D-1和IgY-3D-3均能够选择性且特异性地识别rNTS1,而无法检测rAPJ和rNTS2。值得注意的是,rNTS2和rAPJ与NTS1 ECL2表位区域的氨基酸序列相似性分别仅为36%和27%(补充图8c),这在很大程度上限制了其采用相似空间构象的可能性。
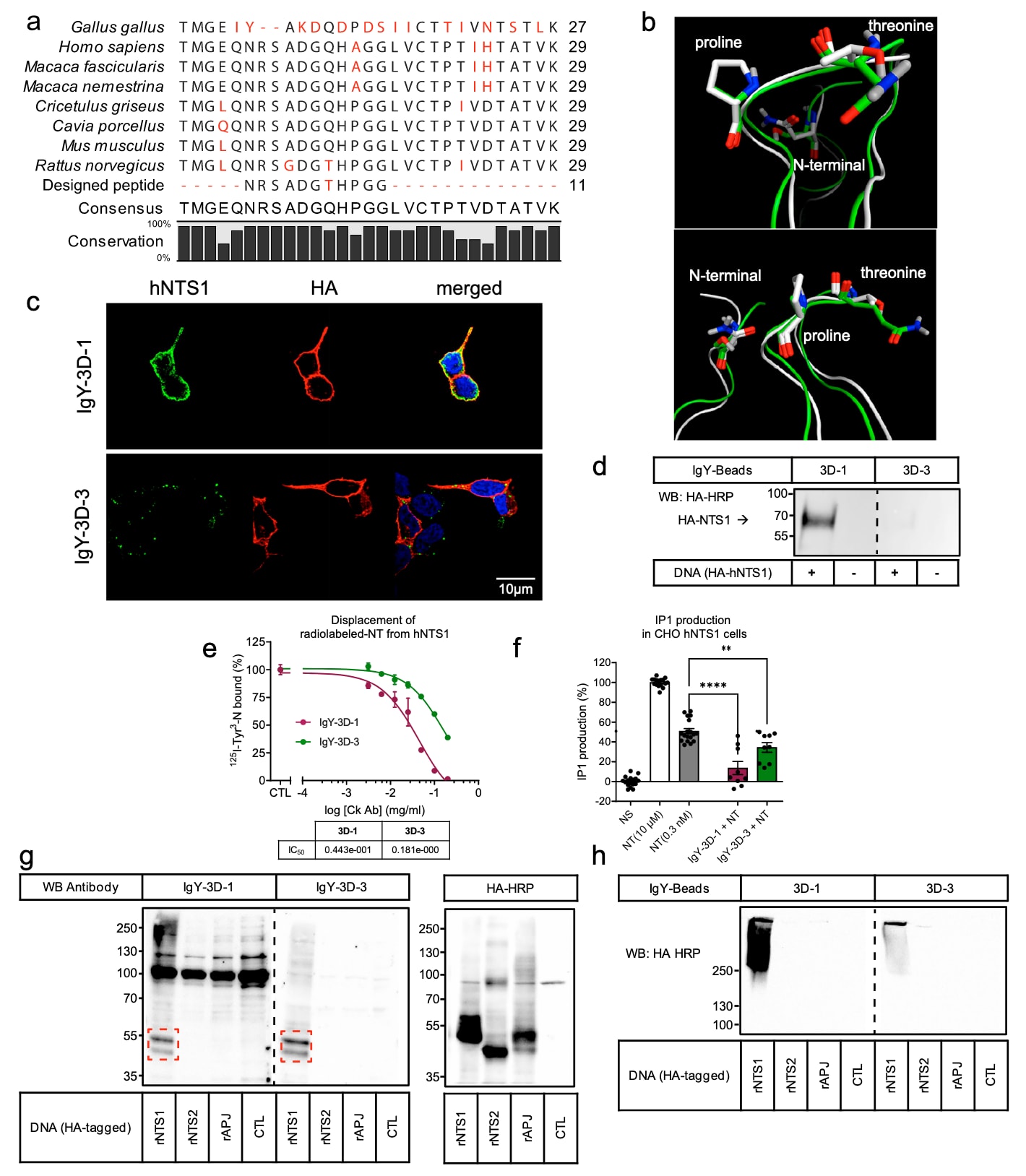
图6 | 抗NTS1抗体的交叉反应性与选择性。 a 不同物种NTS1的ECL2区域对应氨基酸序列比对,黑色条表示序列保守性百分比(CLC sequence bio V5)。b 人源(绿色带状结构)与大鼠(白色带状结构)NTS1的ECL2环结构叠合图,标出了rNTS1与hNTS1之间不同的两个氨基酸(脯氨酸和苏氨酸),并标示了N端结构域。c 稳定表达HA标记人源NTS1受体的固定HEK293A细胞的免疫荧光显微图像。绿色信号表示检测到的hNTS1(IgY-3D-1和IgY-3D-3),红色信号表示HA特异性检测(HA-HRP)。d 使用偶联磁珠的鸡源抗体对稳定表达HA-hNTS1的细胞进行免疫共沉淀,随后采用抗HA抗体进行免疫印迹检测。转染空载体的细胞作为对照。e IgY-3D-1和IgY-3D-3对放射性标记NT的置换曲线,采用三参数非线性回归分析。数据以平均值 ± 标准误表示,每个条件包含3个技术重复,代表两次独立的生物学实验(n = 2)。f IgY抗体对NT诱导的IP1生成的抑制作用。阴性对照为未刺激细胞(NS),所有抗体均在NT(EC50)存在条件下进行检测。数据以平均值 ± 标准误表示,每个实验至少包含9个技术重复,代表两次独立的生物学实验(n = 2)。统计分析采用单因素方差分析并结合Dunnett多重比较检验,P值标注如下:**表示P = 0.0020,****表示P < 0.0001(GraphPad在校正后的精确P值低于软件阈值时报告为<0.0001)。瞬时表达HA标记的大鼠NTS1、大鼠NTS2或大鼠APJ受体的HEK293A细胞分别用于Western blot分析(g)和天然条件下的免疫共沉淀(h),所用抗体为IgY-3D-1或IgY-3D-3。结果代表至少两次独立实验。原始数据已作为Source Data文件提供。
3 讨论
GPCRs为生物治疗药物,尤其是抗体类药物的开发提供了重要机遇。然而,目前获批的绝大多数靶向GPCR的药物仍以小分子或肽类为主。相较之下,抗体因其更优越的药代动力学和药效学特性,包括更有利的体内分布以及更长的半衰期,从而可减少给药频率,同时具备更高的特异性,因而不易引发脱靶毒性。抗GPCR抗体开发进展缓慢的主要原因在于GPCR本身免疫原性较弱、哺乳动物物种间序列高度保守,以及难以制备保持天然构象活性的GPCR抗原。迄今为止,尚无策略能够稳定可靠地产生模拟目标GPCR天然结构的表位。
为克服这些挑战,该研究提出并验证了一种新策略,利用模拟GPCR胞外环三维结构的宏环肽作为免疫原,并以鸡作为免疫宿主来制备抗原特异性抗体。由于鸡与哺乳动物之间存在约2亿年的系统发育差异,这一策略有助于增强对保守哺乳动物蛋白(如GPCR)的免疫反应,从而获得高亲和力、高滴度的抗体。这一优势已在其他针对高难度靶点(包括GPCR)的研究中得到验证。此外,鸡源抗体可通过多种策略实现嵌合化或人源化,例如结合CDR移植和基于人工智能的计算预测工具,以选择最合适的人类生殖系序列,从而实现高效转化。
该研究进一步支持了使用三维表位而非传统N端或C端线性表位来靶向GPCR的有效性。通过分子建模与预测分析软件进行的宏环表位模拟物与连接臂的理性设计,是获得高特异性和高选择性鸡源空间抗GPCR抗体的关键步骤。该研究选择NTS1作为靶点,该受体是一个研究较为深入的A类GPCR,在包括结直肠癌和非小细胞肺癌在内的多种人类癌症中高表达。NTS1的高表达还与肿瘤侵袭性增强及不良预后密切相关,尤其是在胰腺腺癌、三阴性乳腺癌和胶质母细胞瘤中,突显了其作为潜在预后和治疗标志物的重要性。此外,NTS1与NT形成复合物的晶体结构的解析,进一步强化了表位三维结构理性设计的科学基础。
该研究中用于生成抗NTS1抗体的3D抗原肽的关键特征及相应IgY抗体的性质总结于表2。研究结果表明,IgY-3D-1和IgY-3D-3均能够特异性识别NTS1及其三维结构,其中IgY-3D-3还表现出对NT结合状态下NTS1受体的拮抗活性,这一特性在治疗应用中尤为引人关注,尤其是在目前尚缺乏仅对NTS1具有高度选择性且不与NTS2交叉反应的小分子拮抗剂的背景下。
表2 | 三种3D肽及其对应IgY抗体主要特征的总结
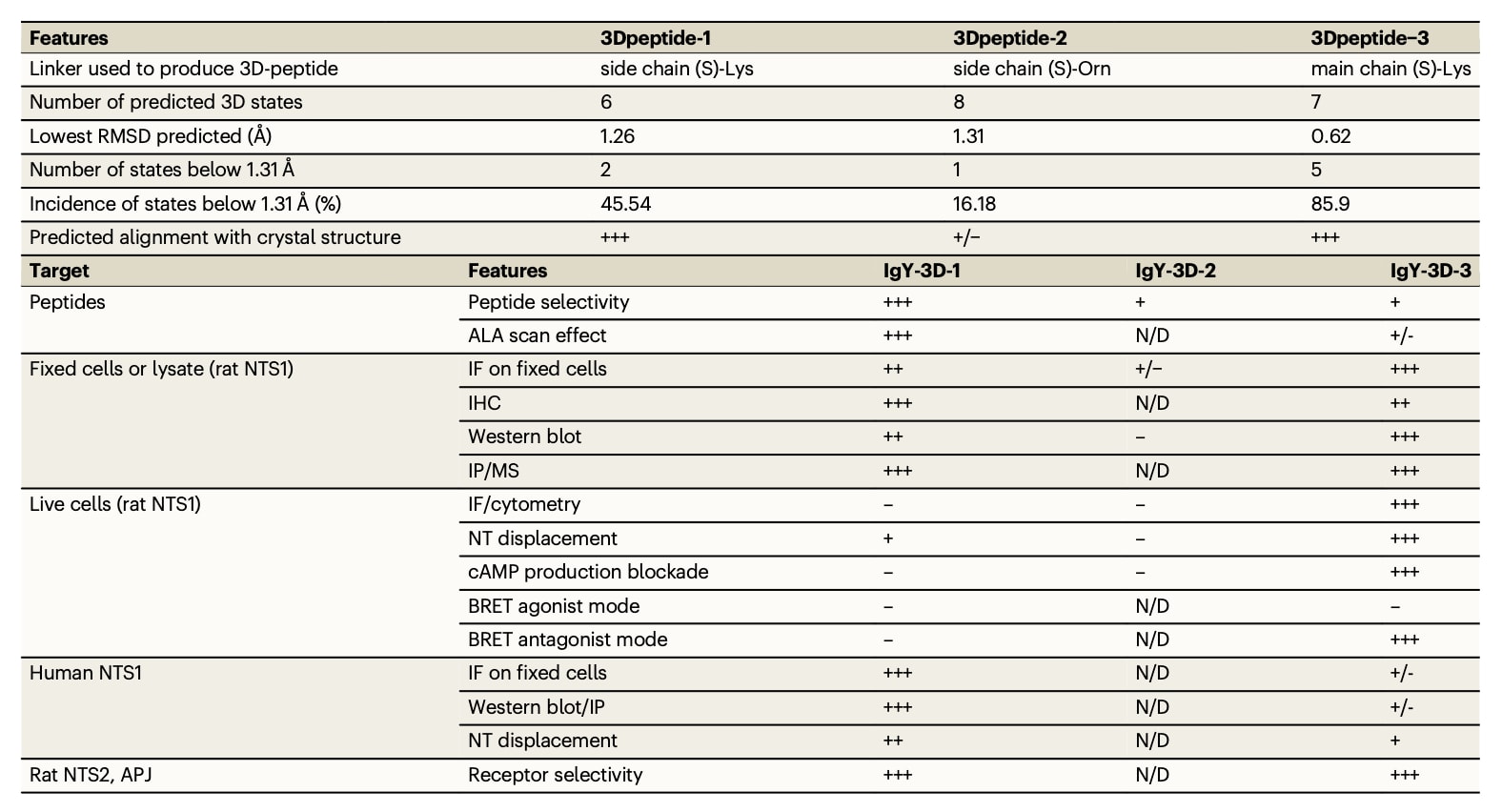
抗NTS1 3D抗体的实验结果与分子建模分析及设计肽的三维结构预测高度一致。与(S)-鸟氨酸连接臂(3Dpeptide-2)相比,(S)-赖氨酸连接臂(3Dpeptide-1和3Dpeptide-3)在鸡体内表现出更高的靶点亲和力,这一点也得到了ECL2模拟分析的预测支持。IgY-3D-2无法检测NTS1,与其预测构象中绝大多数状态的RMSD值超过1.31 Å且仅有16.18%的构象位于阈值以内密切相关。连接臂的重要性还体现在两种宏环肽在成环方式上的细微差异,即侧链成环的3Dpeptide-1与主链成环的3Dpeptide-3,这一差异直接决定了抗体在与NT结合状态下是否具备拮抗受体活性的能力。3Dpeptide-3中低于1.31 Å阈值的构象状态多达5种,累计占比达85.9%,明显高于3Dpeptide-1的45.54%,这可能解释了由3Dpeptide-3诱导产生的多克隆抗体所具备的额外功能特性。
值得强调的是,IgY-3D-1和IgY-3D-3均对NTS1具有高度选择性,无法识别与其高度相关的NTS2或同属GPCR家族的APJ受体。总体而言,这一概念验证研究充分展示了计算模拟技术在宏环表位设计中的有效性与价值。随着多种具有治疗意义的GPCR结构不断被解析,为空间IgY抗体的设计提供了重要契机。
这些鸡源抗NTS1抗体依据抗体验证指南进行了系统而严格的验证,在ELISA、固定与活细胞免疫荧光显微镜、免疫组织化学、流式细胞术、Western blot、免疫共沉淀以及质谱等多种分子和细胞实验中均表现出良好的特异性与选择性。更重要的是,通过放射性配体结合实验和基于BRET的活细胞生物传感器分析,IgY-3D-3被证实在NT激活NTS1后,可作为G蛋白和β-arrestin信号通路的拮抗剂发挥作用。此外,通过检测不同批次抗体,证实其在特异性识别NTS1方面具有良好的重复性,有效缓解了抗体批次差异这一长期存在的问题。
当然,多克隆抗体研究本身也存在固有局限性,其观测到的特征往往来源于多种免疫球蛋白亚群的综合效应,因此结果反映的是整体趋势而非单一分子的精细特性。此外,免疫纯化过程可能导致部分高亲和力结合子丢失,从而影响免疫反应的完整呈现。尽管如此,多克隆抗体在鉴定关键抗原肽段和功能结构域方面仍具有不可替代的价值,尤其是在与计算建模方法相结合时,可显著提升抗原选择的合理性,为功能相关抗体的开发提供战略优势。一旦确定最优抗原肽,即可借助噬菌体展示、酵母展示或单B细胞筛选等先进技术,更加高效、低风险地开展单克隆抗体的发现工作。初始文库可来源于免疫鸡的外周血单核细胞、冷冻保存的脾组织或骨髓样本,从而通过迭代筛选获得具有理想表型和功能的克隆。
综上所述,该研究成功制备并验证了两种可靠的靶向NTS1的抗GPCR抗体。其中IgY-3D-3展现出明确的功能活性,在治疗应用方面具有潜在价值,而IgY-3D-1则表现出高度选择性,更适合作为诊断工具。除传统的治疗和诊断用途外,靶向GPCR的抗体还可进一步设计以赋予效应功能,例如通过抗体依赖的细胞毒作用(ADCC)清除肿瘤细胞,或构建抗体药物偶联物(ADC),以及能够同时结合两种不同抗原或表位的双特异性抗体,从而调控GPCR的同源或异源二聚化。最后,由于抗体能够实现对GPCR不同构象状态的识别,其在开发具有功能选择性的治疗策略方面具有广阔前景。总体来看,该空间抗体生成策略为GPCR相关疾病,尤其是癌症、感染性疾病及代谢性疾病的诊断和治疗提供了重要而有前景的研究方向。