JACS 2026 | 优化动态小分子结合蛋白的稳定性
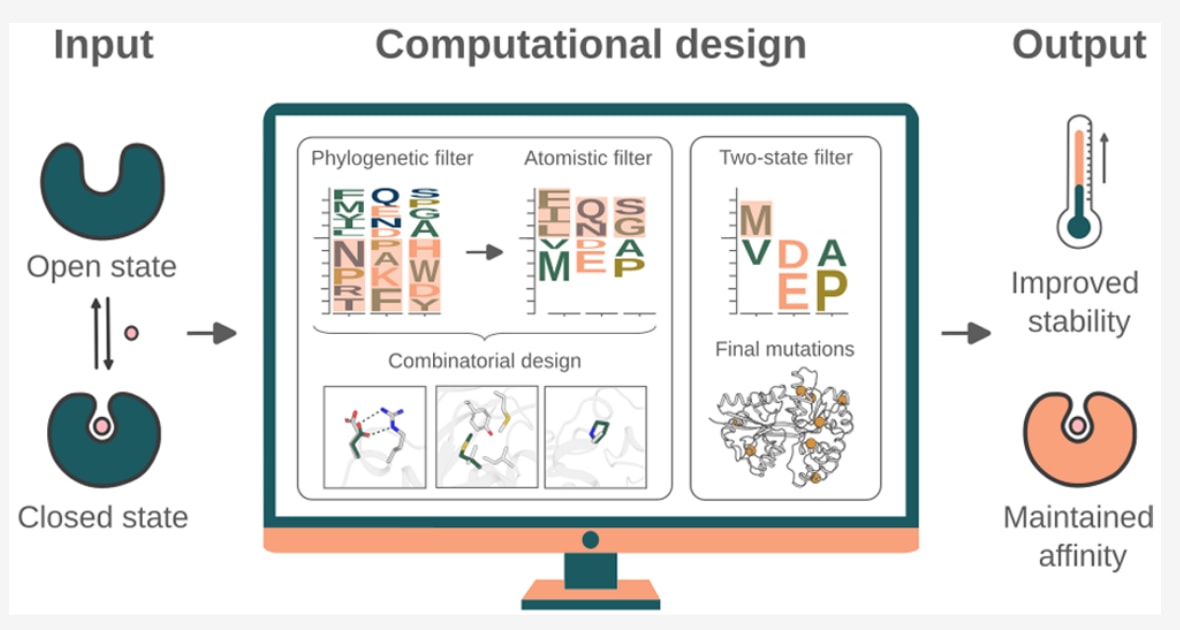
动态蛋白在药物发现与生物传感中扮演关键角色,但其在不同构象状态之间频繁切换,使稳定性设计长期面临“提稳即失活”的难题。发表在JACS的这项研究聚焦于动态小分子结合蛋白,系统揭示了单一构象稳定化策略在动态体系中的内在局限。作者以周质结合蛋白这一经典动态蛋白超家族为模型,比较了开放态与闭合态下突变能量景观的显著差异,指出仅基于单态和进化约束的设计往往破坏配体结合能力。通过引入双构象一致性筛选,并对铰链和界面区域施加结构约束,研究成功在较低突变负载下显著提升蛋白热稳定性,同时保持近似野生型的结合亲和力。该工作不仅为动态蛋白的理性稳定化提供了清晰可行的设计范式,也为高稳定性、生物响应型传感器蛋白的工程化奠定了重要基础。

0 摘要
动态蛋白质的功能由其不同构象状态的稳定性以及分隔这些状态的能垒所决定。对于大多数动态蛋白而言,这些能垒的分子层面细节尚不清楚,这意味着现有蛋白设计方法在不破坏活性的前提下引入有益突变方面存在根本性的限制。基于此,研究提出一种假设,即设计能够同时兼容结构上显著不同的平衡构象的突变,可能实现更可靠的稳定性设计。
研究以周质结合蛋白作为研究对象,这是一类高度动态的蛋白超家族,在与小分子配体结合时会从开放态转变为闭合态。结果表明,针对一种构象计算得到的、受进化约束的允许突变空间,与另一种构象对应的突变空间并不相容。因此,在设计过程中进一步对潜在的构象铰链位点和界面残基施加约束,并过滤掉彼此不兼容的突变。
以四种不同的周质结合蛋白为起点,共设计获得了16个稳定化变体,每个变体包含7至28个突变。结果显示,仅基于单一构象并结合进化约束进行的设计,无法维持类似野生型的配体结合亲和力。相反,选取同时兼容两种构象并满足结构约束的一部分突变,可以在显著提高热稳定性的同时,最大程度减轻对配体结合能力的负面影响。该研究展示了一种直接而有效的方法,可实现动态蛋白的一步稳定化,这对于获得具有高热稳定性和良好响应性的生物传感器起始蛋白至关重要。
1 引言
许多蛋白质具有动态特性,能够在平衡条件下或在结合其他分子时在多种构象状态之间切换。蛋白质动力学使其在功能上不同的状态之间发生转变,从而实现变构调控和协同效应等复杂功能。动态蛋白的折叠能量景观可能在进化过程中被精细调节,既能够促进功能状态之间在纳秒至秒尺度内的快速交换,又避免蛋白质被锁定在单一构象中。尽管这些特性至关重要,决定蛋白构象平衡的能量景观依然十分复杂,目前理解有限,且除少数小型、快速折叠的蛋白外,难以通过计算手段准确估计。
该研究聚焦于提升动态结合蛋白的稳定性。对于构象变化较小的蛋白,将系统发育分析与单一构象上的能量计算相结合的方法,例如PROSS和FireProt,已被证明具有较高的可靠性和广泛适用性。然而,当这些方法应用于动态蛋白时,稳定性设计的结果并不一致。例如,基于单一配体和共激活因子结合晶体结构,PROSS成功提高了人雌激素受体α的稳定性、表达性及结晶能力,且未观察到明显的活性下降;类似地,该方法也改善了电压门控钾通道的表达性。在这两种情况下,亚基界面和潜在铰链位点均被限制为不可突变。相反,在未考虑铰链位点的情况下,将PROSS应用于周质结合蛋白DalS和氨基酯水解酶QVH时,却导致蛋白活性下降。既往的设计研究还表明,周质结合蛋白在计算设计中可能是异常具有挑战性的对象,常出现未预期的构象变化、稳定性降低以及不理想的功能表现。
细菌周质结合蛋白是一类多样化的超家族,负责在细菌周质中捕获小分子并将其跨越质膜转运至细胞质。选择周质结合蛋白作为研究对象,一方面源于提升动态蛋白稳定性的普遍需求,另一方面也考虑到此前设计研究中暴露出的具体困难,以及这类蛋白在生物传感领域中的广泛潜在应用。周质结合蛋白通常具有由铰链区域连接的双叶结构,并通过类似捕蝇草的机制在开放态与闭合态之间切换以结合不同配体。尽管整体折叠方式相似,其序列却高度多样,相应的能量景观也十分复杂,包含多个亚稳态。
值得注意的是,已有多种策略将周质结合蛋白改造为基于荧光的生物传感器,主要用于生物医学研究。然而,引入荧光蛋白结构域或优化配体特异性与结合亲和力,往往伴随着表达产量下降和热稳定性降低。因此,一种可靠的周质结合蛋白稳定化策略对于生物传感器工程尤为重要。该研究表明,将稳定化突变限制为同时兼容开放态和闭合态、并避开潜在铰链位点的突变,是一种有效的周质结合蛋白设计方法。由此提出了一种优化动态蛋白稳定性的策略,并为基于周质结合蛋白的生物传感器工程提供了合适的起始蛋白。
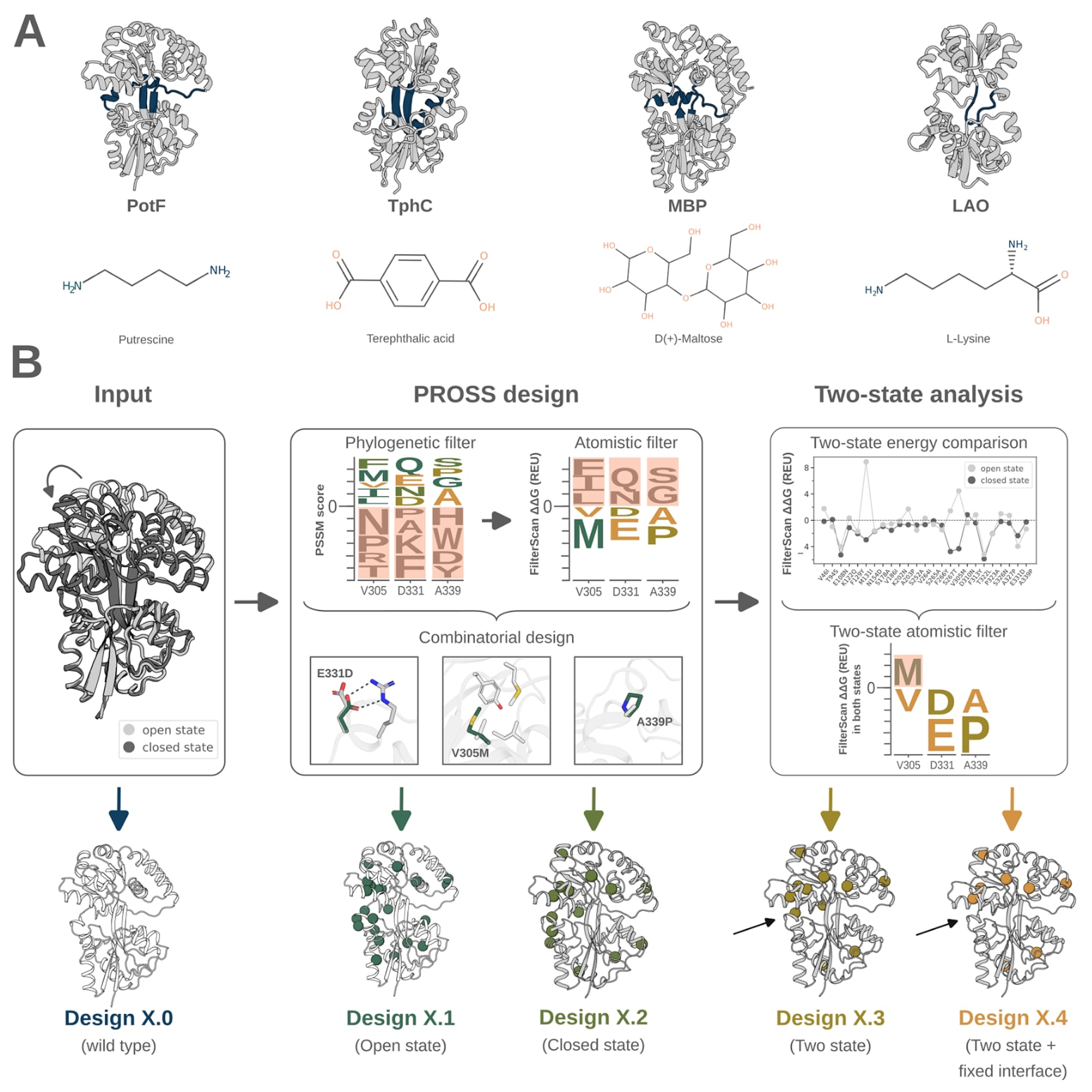
图1|基于单态和双态PROSS设计优化周质结合蛋白稳定性的设计策略。 (A) 选取了四种周质结合蛋白:腐胺结合蛋白PotF、对苯二甲酸结合蛋白TphC、麦芽糖结合蛋白MBP以及赖氨酸结合蛋白LAO。图中以示意图形式展示了闭合态晶体结构(灰色),其中铰链区域以蓝色标出。(B) 设计策略以PotF、TphC、MBP和LAO的开放态与闭合态晶体结构作为输入。所有周质结合蛋白的野生型序列作为对照(设计X.0)。在默认参数下运行PROSS设计计算,分别基于开放态和闭合态结构生成设计X.1和X.2。插图中以PotF的三个PROSS突变为例,展示了新氢键的形成、核心堆积的改善以及环区刚性的增强。突变对能量(ΔΔG)的影响依赖于两种构象状态下的主链构象差异,因此将在任一状态中表现为不稳定(正能量)的突变予以剔除,得到设计X.3。在生成设计X.4时,在运行PROSS设计计算和双态能量筛选之前,对铰链区域和叶片界面进行了固定。设计X.3和设计X.4中的黑色箭头标示了一处由于铰链区域被固定而未出现在设计X.4中的突变。
2 结果
2.1 构象对稳定性设计选择的影响
研究首先检索结构数据库,筛选在配体结合时发生显著构象变化,且开放态与闭合态均已有结构解析的周质结合蛋白。最终选取了四种结合不同类型配体的周质结合蛋白,分别为PotF、TphC、MBP和LAO,其配体涵盖芳香族二羧酸到糖类等多种分子。作为参考,在每一种周质结合蛋白的开放态和闭合态结构上,采用标准参数进行了PROSS稳定性设计计算。随后比较了开放态与闭合态设计中所有突变的ΔΔG值,结果表明,部分突变在一种构象中具有稳定作用,而在另一种构象中则表现为不稳定。总体而言,开放态的能量往往低于闭合态,这可能与闭合态更高的结构堆积密度有关。对这些在不同构象中能量符号发生变化的突变位点进行结构分析发现,其周围主链和侧链构象存在细微差异。
进一步将突变位点划分为表面层、边界层和核心层后发现,表面突变在两种构象中的能量变化相似,可能反映出由于堆积密度较低而对构象变化不敏感。相比之下,边界层和核心层突变,尤其是在PotF和TphC中,表现出明显的构象依赖性能量差异。因此可以认为,蛋白核心区域的突变对构象状态尤为敏感。基于此,研究又生成了两组额外设计方案。第三组设计中,为避免改变周质结合蛋白的构象平衡,排除了在不同构象中能量符号相反的突变,但此时界面区和铰链区仍然存在突变。鉴于这些区域在高斯网络模型分析中显示出最显著的局部主链密度变化,第四组设计进一步去除了铰链区域和叶片界面的突变。最终共获得16个周质结合蛋白设计变体,每种野生型蛋白对应4个设计方案,并与各自的野生型进行实验比较。
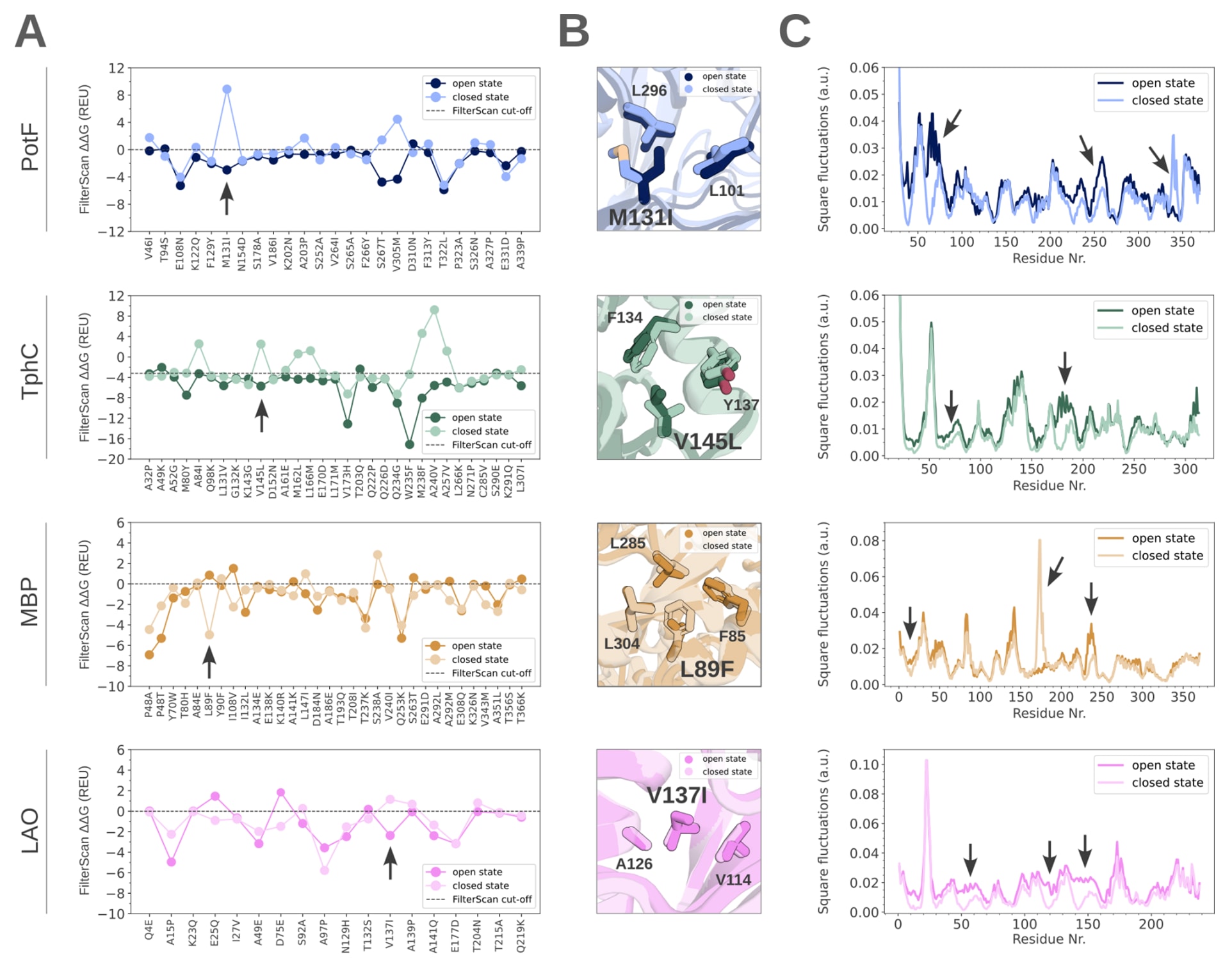
图2|周质结合蛋白单态PROSS突变的Rosetta ΔΔG与堆积密度分析。 (A) 基于PotF(蓝色)、TphC(绿色)、MBP(橙色)和LAO(粉色)的开放态与闭合态结构生成的PROSS突变的ΔΔG值,以及能量截断值(虚线)。能量以Rosetta能量单位表示。箭头指示了在(B)中进行可视化的突变。(B) 在PotF、TphC、MBP和LAO设计中,围绕四个代表性突变位点(以棒状表示)观察到残基堆积的细微差异。这些突变在(A)的分析中表现为在一种构象中具有稳定的ΔΔG值,而在另一种构象中则表现为不稳定。(C) 基于高斯网络模型计算得到的PotF(蓝色)、TphC(绿色)、MBP(橙色)和LAO(粉色)主链平方涨落,其倒数反映了堆积密度。箭头标示了铰链区域和界面残基中堆积密度变化较大的位置。
2.2 双态筛选与结构约束实现可靠的稳定性优化
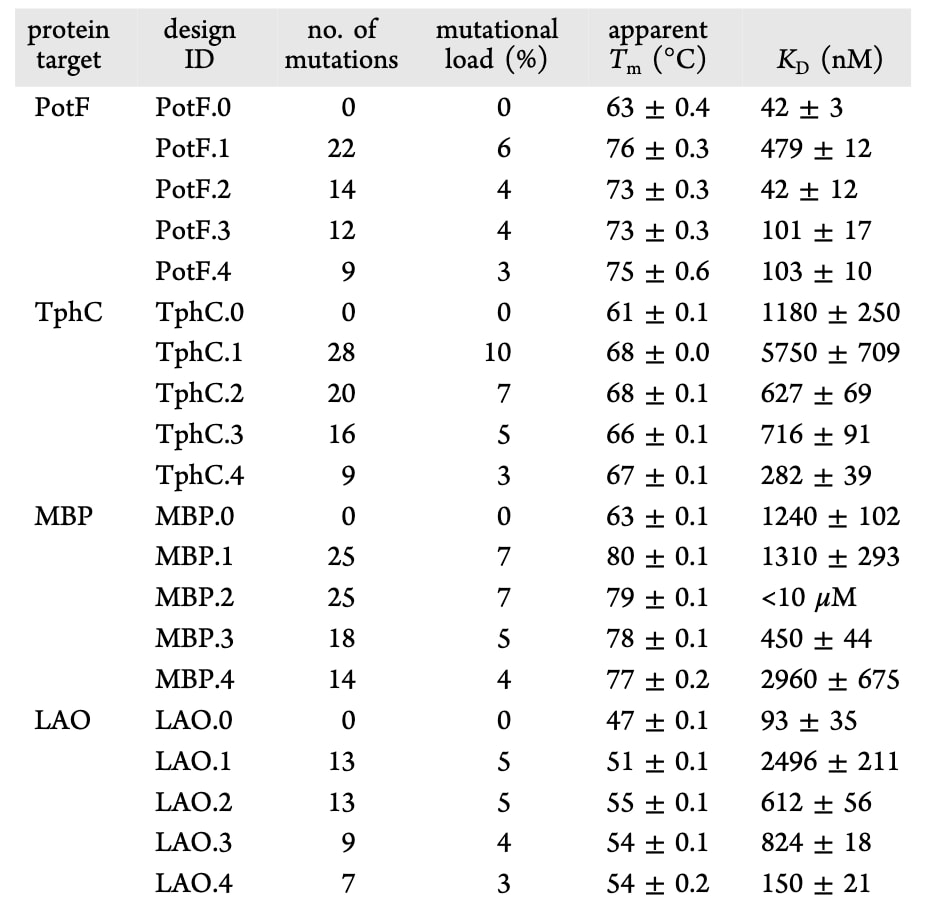
野生型蛋白及其设计变体均在大肠杆菌细胞质中表达并纯化。首先利用圆二色谱检测了所有设计变体的折叠情况,结果表明其整体折叠良好。随后进行的热变性实验显示,所有周质结合蛋白设计变体的热稳定性均得到提升,表观ΔTm介于4至17℃之间。作为对比,野生型PotF、TphC、MBP和LAO的表观Tm分别为63℃、61℃、63℃和47℃。值得注意的是,在PotF、TphC和LAO中,表观Tm的提升幅度与突变数量并不相关,这一现象与以往研究中观察到的趋势不同。
接下来考察了PROSS设计是否仍能结合各自的天然配体。在含有100 μM相应配体的条件下进行了热变性实验,其中PotF、TphC、MBP和LAO分别对应腐胺、对苯二甲酸、麦芽糖和L-赖氨酸。以ΔTm增加1℃作为判断标准,除PotF.4外,所有设计变体均能够结合其天然配体。除三个TphC设计外,大多数周质结合蛋白设计变体的ΔTm值略低于野生型蛋白,提示其配体结合亲和力可能有所下降。
为更深入分析突变对配体结合行为的影响,进一步采用等温滴定量热法测定了设计变体的结合常数,并与对应野生型蛋白进行比较。野生型PotF、TphC、MBP和LAO的结合亲和力分别为42±3 nM、1180±250 nM、1240±102 nM和93±35 nM,与已报道的文献数值基本一致。在单一构象设计中,8个变体中有3个的结合亲和力降低超过5倍,其中2个基于开放态构象设计。对于基于闭合态的MBP.2,其等温滴定量热数据无法得到可靠拟合。相比之下,未施加铰链约束的双态筛选设计仅在4个案例中有1个出现亲和力下降,而所有同时施加双态筛选和铰链约束的设计均保持了与野生型相近的结合亲和力。在TphC.2–4和MBP.3中,结合亲和力甚至出现了轻微提升。综合来看,仅基于单一构象进行周质结合蛋白设计更容易改变其配体结合亲和力,而结合双态突变筛选,尤其是对铰链区域施加约束,能够在可靠提升热稳定性的同时保持配体结合能力。
表1|周质结合蛋白设计变体及野生型的热稳定性和配体结合参数。
3 讨论
在缺乏对底层能量景观充分认识的情况下优化动态蛋白是一项极具挑战性的任务,因为构象状态与功能通常紧密耦合。热变性和配体结合实验结果表明,在周质结合蛋白的单一构象设计中,仅依赖进化约束并不足以维持类似野生型的结合亲和力。要在快速时间尺度内实现开放态向闭合态的转换,同时又稳定开放态构象集合以便配体进入,可能需要精细的能量平衡。通过突变能量比较揭示的开放态与闭合态之间堆积密度差异,可能正参与了这种构象平衡的维持。位于结合口袋的突变能够影响周质结合蛋白的构象分布以及闭合态的可达性,而铰链区域的突变则可能通过差异性稳定特定主链构象而削弱配体结合能力。研究结果还显示,即便是远离功能区域的突变,也可能在不同构象中表现出能量符号的差异。
利用第二种构象的结构信息提高了设计方法的可靠性,但在LAO.3中仍观察到结合亲和力下降。因此,只有在进一步对铰链和界面残基施加约束后,才能在所有案例中实现成功设计,即在显著减少突变数量的情况下,仍获得与高突变负载设计相当的热稳定性提升。由此可见,该策略能够在较低突变负载下实现周质结合蛋白的可靠优化,并为生物传感器工程提供稳健的起始蛋白。
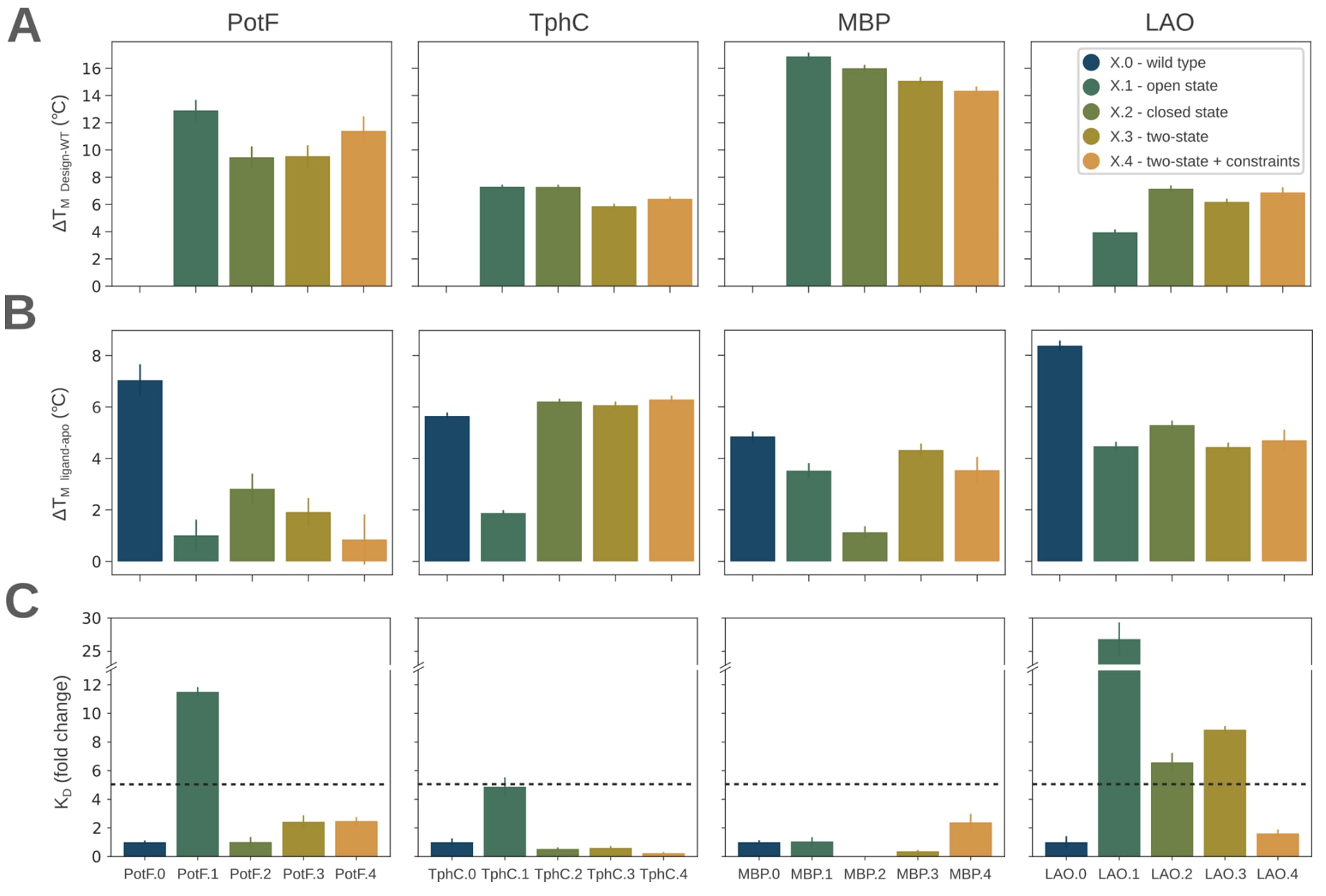
图3|周质结合蛋白野生型及其设计变体的热稳定性和结合亲和力。 (A) 表观熔解温度变化ΔTm相对于野生型计算。(B) 在加入100 μM配体后表观ΔTm的变化,通过减去相应无配体对照样品得到。(C) 相对于野生型的结合常数KD变化倍数。误差线表示三次独立重复实验的标准偏差。在PotF、TphC、MBP和LAO的结合实验中,分别使用腐胺、对苯二甲酸、麦芽糖和L-赖氨酸作为配体。相对于野生型结合亲和力降低5倍的位置以虚线标示。
4 总结
尽管该设计策略在四种周质结合蛋白中表现良好,其是否能够推广至其他动态蛋白仍有待验证。对于大多数动态蛋白而言,目前尚缺乏覆盖所有功能状态的高质量结构信息。若能够借助AlphaFold等基于机器学习的方法,高精度地构建动态蛋白的结构集合,该设计策略的适用范围有望进一步拓展。该研究中发现,AlphaFold3在默认设置下生成的模型中,至少有五分之一能够近似再现所研究周质结合蛋白野生型的开放态和闭合态构象。然而,仍需确保所选用的结构确实代表蛋白的功能状态,否则对非功能构象的稳定化将对功能产生不利影响。总体而言,该研究提出了一种用于稳定化动态蛋白的设计方法,并为理解和设计复杂动态蛋白迈出了重要一步。