Nat. Chem. Biol. 2025 | 利用深度学习对三个不重叠的蛋白质表位进行精确的单域支架构建
今天介绍的这项工作来自 Nature Chemical Biology。天然蛋白通常在单一结构中承担多种功能,而从头设计的蛋白大多只能精确呈现单一功能位点,核心限制在于可设计结构空间以及多表位之间复杂的空间约束关系。该研究围绕这一长期难题,提出了一种基于深度学习的多表位支架设计框架,在不预设各表位刚体取向的情况下,于单一小型结构域蛋白中同时精确嵌入三个彼此不重叠的病毒中和表位。该研究以呼吸道合胞病毒融合蛋白为模型,设计的免疫原长度不足130个氨基酸,近一半表面积由表位构成,其整体折叠显著偏离已知蛋白结构数据库中的天然拓扑。X射线晶体学结果表明,三个位点在主链构象与关键侧链取向上均被高保真复现。在动物免疫实验中,多表位设计相比单表位免疫原显著提高了对天然病毒蛋白的交叉反应抗体滴度,并诱导出具有生理相关性的中和活性。该工作系统展示了生成式深度学习在高度受限蛋白设计问题中的优势,不仅为单组分替代多组分鸡尾酒疫苗提供了新范式,也为多功能蛋白、治疗性分子及人工酶的理性设计拓展了结构空间与方法学边界。

获取详情及资源:
- 📄 论文: https://www.nature.com/articles/s41589-025-02083-z
- 💻 代码: https://github.com/RosettaCommons/RFjoint2.git
0 摘要
从头蛋白设计在单一功能基序的支架化方面已经取得了显著成功,然而在天然体系中,大多数蛋白质往往同时呈现多个功能位点。该研究提出了一种利用深度学习在单一结构域蛋白中同时支架化多个功能位点的方法。该研究设计了一类长度不足130个氨基酸的小型单结构域免疫原,在同一蛋白中呈现来源于呼吸道合胞病毒的三种彼此不同且结构不规则的基序。这些基序合计占据了设计蛋白近一半的序列与表面积,因此其整体折叠形式十分新颖,与蛋白质数据库中已知结构几乎不存在全局相似性。尽管如此,X射线晶体结构分析证实,各个基序均以高精度被正确呈现;与单表位免疫原相比,多表位设计能够诱导更高水平的交叉反应抗体滴度和更强的中和反应。在一个小型单结构域蛋白中成功呈现三种不同的结合界面,充分体现了生成式深度学习方法在解决复杂蛋白设计问题方面的强大能力。
1 引言
近年来,计算蛋白设计领域的快速发展使得研究者能够以原子级精度生成种类极其丰富的蛋白序列与结构,并在实验层面不断取得成功。相比之下,传统计算方法在嫁接复杂结构基序时,往往受限于少数已被充分表征、能够容纳此类基序的天然蛋白结构,这在很大程度上限制了为设计蛋白赋予功能的能力。因此,迄今为止,大多数从头设计蛋白仍然局限于通过单一支架化基序实现单一功能。与之形成鲜明对比的是,天然蛋白通常具有多重功能,这些功能往往源于基因融合等进化过程。随着神经网络在结构预测以及结构或序列生成方面的进展,该研究者得以在天然蛋白空间之外探索并设计多样化的全新折叠形式。除能够采样复杂拓扑结构之外,基于深度学习的工具还展示了生成嵌入生物学功能的合理支架结构的能力。这类方法在很大程度上打破了对既有蛋白结构空间的依赖,并有望提供将多个不同基序嵌入定制化从头支架所需的构象灵活性。
为从头设计蛋白赋予多个功能基序,将显著拓展其潜在应用范围。在疫苗学领域,单一抗原同时诱导针对多个表位的免疫应答,有望推动更优免疫原的设计。通过在设计蛋白上展示病毒表位,可以有针对性地呈现中和表位,从而触发特异性的免疫反应,同时避免那些免疫优势更强但不具中和作用的位点,提高抗体反应的效率。所谓反向疫苗学正是试图将免疫反应引导至预先定义的抗原决定簇,但病毒表位通常呈现为不规则且不连续的结构,使其在从头设计蛋白上的精确复现成为一项极具挑战性的任务。已有研究表明,支架化的免疫表位能够成功诱导针对目标位点的抗体反应,并且以鸡尾酒疫苗的形式可以重塑整体免疫应答。然而,利用从头设计的免疫原在初次免疫动物中诱导高滴度抗体仍然是一个悬而未决的问题,其中一个重要原因在于免疫原表面相当大的比例被支架蛋白本身占据,而非真正具有免疫价值的表位。若能够在同一支架上呈现多个彼此不重叠的表位,将在很大程度上减少不必要的支架表面积,从而有望提升免疫原的整体效果。
在该研究中,基于深度学习的方法被用于设计嵌入多达三个功能位点的从头蛋白支架。通过在新生成的支架蛋白上同时展示呼吸道合胞病毒融合蛋白的三个表位,构建了一个面积更大、组成更为复杂的抗原表面,从而有效减少了不属于表位的支架暴露区域。研究结果表明,这三个免疫表位能够在同一从头设计的支架上被同时复现,并且在结构层面具有很高的精确度,即使这些支架的整体拓扑显著偏离既往表征的蛋白结构空间。进一步的免疫学实验显示,与单表位免疫原相比,多表位设计能够诱导更高水平的针对天然融合蛋白的交叉反应抗体滴度,显示出其作为鸡尾酒疫苗替代方案的潜在价值。更为广泛地看,该研究提出了一种通用的方法学框架,用于在高度未知的蛋白拓扑空间中设计嵌入多种基序的蛋白,为构建能够容纳复杂结构基序和多重结合位点的蛋白分子提供了新的思路。
2 呈现一种新型RSVF表位的从头设计支架蛋白
该研究首先考察了深度学习工具是否能够解决传统设计方法长期难以攻克的单表位支架化问题。呼吸道合胞病毒融合蛋白的V位点表位由一段螺旋–转角–β链构成,由于其与蛋白质数据库中已知结构的相似性极低,无法直接嫁接到天然蛋白支架上。此前的概念验证研究中,曾利用RoseTTAFold的基序约束幻觉方法将该表位支架化到从头设计蛋白中,但仅获得了对V位点特异性抗体RSV90具有微摩尔级亲和力的弱结合体。在此基础上,为获得更高亲和力的设计,该研究扩大了幻觉设计的规模,并引入ProteinMPNN序列设计工具以改善蛋白的稳定性和溶解性。通过酵母表面展示结合筛选,共检测了4547个设计,其中492个在100 nM条件下能够与RSV90结合。在筛选得到的候选中,有39个被选中进行进一步分析,其中27个成功在大肠杆菌中表达,并对结构差异性最大的四个设计体RSVFV-1至RSVFV-4进行了系统表征 (图1a)。结果显示,这些设计在幻觉模型与AlphaFold2预测之间具有良好一致性,圆二色谱表明其呈现混合的αβ二级结构特征,在高达90 ℃条件下几乎不发生解折叠,并且在体积排阻色谱结合多角度光散射分析中主要以单体形式存在。四种支架蛋白与RSV90的解离常数位于54至241 nM之间 (图1b),与天然RSVF三聚体的亲和力相比相差不超过50倍,同时比此前报道的最佳RSVFV单表位设计提高了约20倍。进一步解析了RSVFV-1与RSV90 Fab复合物的晶体结构,分辨率达到2.4 Å。该结构与AlphaFold2预测高度一致,其主链均方根偏差为1.05 Å (图1c),且被支架化的RSVFV表位在主链和侧链构象上均与设计所依据的RSV90结合态RSVF结构高度吻合,基序的主链均方根偏差为0.843 Å,全原子偏差为1.37 Å (图1d,e)。综合这些结果表明,深度学习方法能够以极高的结构精度支架化抗原表位,相比以往的RSVFV单表位设计在性能上取得了显著提升。
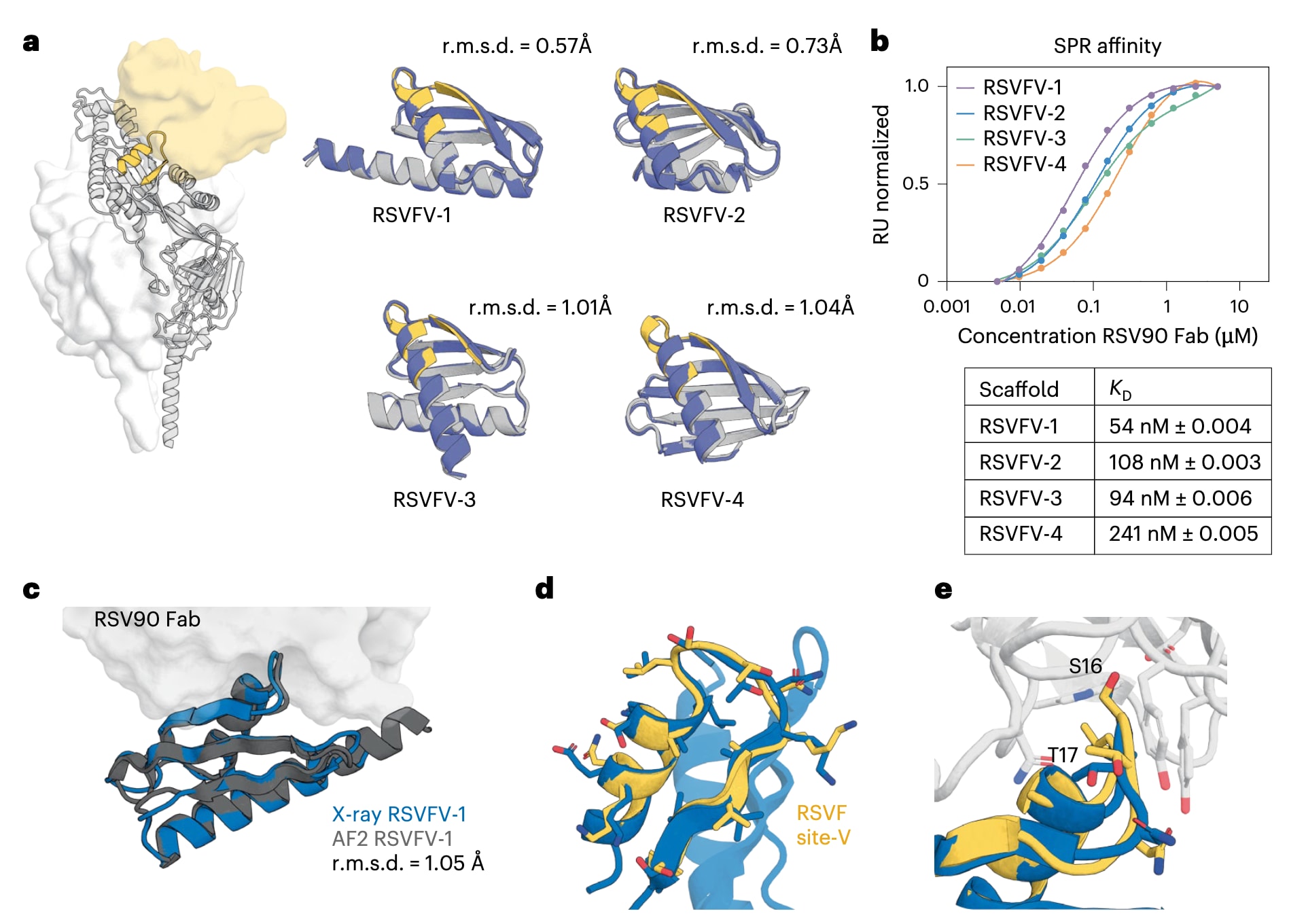
图1|RSVFV免疫原候选体的表征。 a,在RSVF三聚体结构中标示RSVFV表位,并显示其与V位点特异性抗体结合的界面(黄色表面)。左侧为针对RSVF V位点进行支架化的优选候选体,其RoseTTAFold幻觉模型(灰色)与AlphaFold2预测结果(蓝色)叠合展示,并给出了各模型的全原子均方根偏差。b,采用表面等离子体共振技术测量四种设计体与RSV90 Fab之间的稳态结合亲和力,误差为基于曲线拟合计算得到的标准误。c,RSVFV-1与RSV90 Fab形成复合物的晶体结构与AlphaFold2模型高度一致,其主链均方根偏差为
3 嵌入三种RSVF中和表位的从头设计支架蛋白
鉴于基于深度学习的蛋白设计方法在以往被认为难以支架化的表位上已取得实验成功,研究进一步探讨了是否能够在单一支架上同时呈现多个结构复杂程度不同的表位,以期获得表位与支架表面积比例更高、抗原谱更广的免疫原。尽管已有计算设计方法成功将功能基序嵌入从头设计的支架中,但针对在非天然取向下于单结构域蛋白中呈现多个不同表位的研究仍然十分有限。
为在同一支架中嵌入三种已知可诱导中和抗体的RSVF表位 (图2a),该研究采用了基于深度学习的RFjoint2 Inpainting方法,该方法源自RoseTTAFold结构预测网络。在设计过程中,将RSVF的II、IV和V位点表位片段作为模板输入网络,但并未预先限定它们之间的相对取向(图2b)。由于这些表位在结构上彼此独立,且在很大程度上是自洽的,其刚体取向并非诱导抗体应答所必需,这种取向上的自由度显著拓展了可生成支架的多样性。在完成RFjoint2 Inpainting生成骨架之后,该进一步利用ProteinMPNN进行序列设计,并按照既往流程通过AlphaFold2进行筛选 (图3a)。为避免抗体在同一支架上同时结合时发生空间位阻,将相应抗体对齐至设计支架上的表位,并剔除了在抗体结合模式上存在结构不相容的设计(图3b)。由于未预设表位的刚体位置,RFjoint2得以采样到范围广泛的表位空间排布方式。
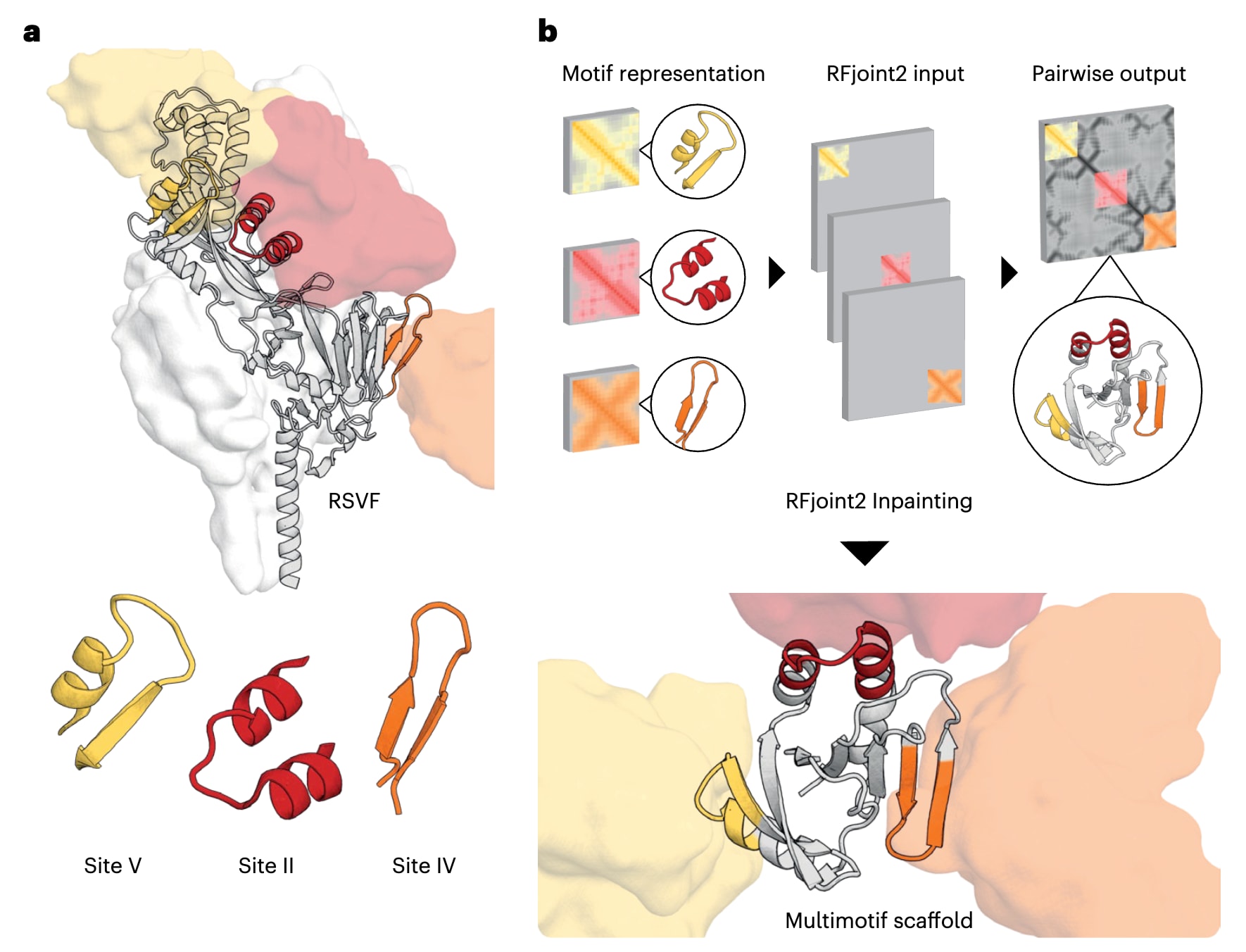
图2|单表位与多表位免疫原的计算支架化设计。 a,在RSVF结构中标示用于单表位和多表位免疫原设计的目标表位,同时展示针对V位点、IV位点和II位点的抗体可变区,其结构分别对应PDB 5TPN、PDB 3O45和PDB 3IXT,不同表位以相应颜色的表面表示。b,多基序支架化设计中,将各个表位基序分别作为模板输入RFjoint2。由于模板输入对整体三维空间位置不敏感,因此在为每个基序提供独立模板时,三种基序之间的相对空间位置并未被预先限定;相反,RFjoint2在设计过程中自动构建一个支架蛋白(灰色),以非天然的相对取向同时支撑这三种基序(彩色)。
在筛选后的多表位设计体中,该研究选取了32条序列进行酵母表面展示筛选。为识别能够同时正确复现三种表位的折叠良好支架,首先通过有限蛋白酶消化并结合II位点特异性抗体进行初筛,随后依次使用IV位点特异性抗体和V位点特异性抗体进行两轮筛选。经过三步筛选,从32个序列中获得了5条高频重复序列,其中4条在细菌中表现出良好的表达水平。对4个多表位设计体进行圆二色谱分析显示,其二级结构特征与设计模型一致,呈现混合的αβ构象,在高达90 ℃条件下具有较高的热稳定性,并且在体积排阻色谱中主要以单体形式存在 (图3c)。四种支架蛋白均能够与三种位点特异性抗体结合。结合亲和力定量分析表明,II位点和IV位点特异性抗体对所有设计体的结合均处于纳摩尔量级,其中II位点抗体的亲和力为14至47 nM,IV位点抗体为343至890 nM,相比之下,天然RSVF三聚体对相应抗体的亲和力更高,这可能反映了在同一支架中整合三种表位所带来的设计难度(图3e-g)。相对于天然RSVF三聚体,设计体对V位点特异性抗体的亲和力较低,可能源于该表位本身更高的结构复杂性 (图3e)。总体而言,这些结果首次表明,能够同时呈现多个功能位点的从头设计蛋白可以以相对较高的实验成功率被构建出来。
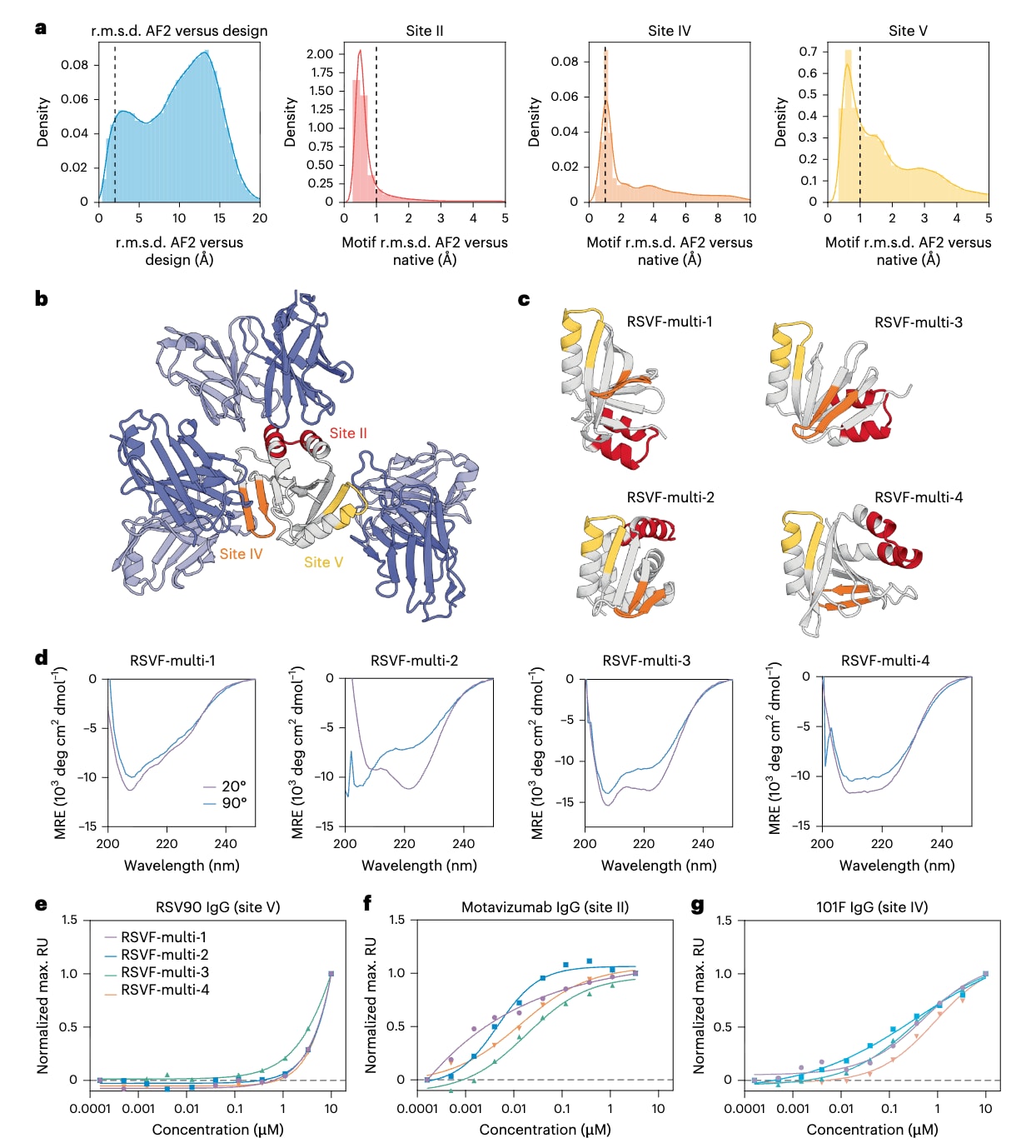
图3|多表位免疫原候选体的实验表征。 a,计算层面的评估,利用AlphaFold2评估预测结构与设计模型之间的一致性,以及预测结构中各表位与天然表位结构之间的相似程度,图中的竖线表示计算筛选中判定为“成功”的阈值。b,RFjoint2生成的代表性设计示例,尽管在设计过程中并未显式约束抗体可及性,但各个表位仍保持对其目标抗体的可结合性。图中将RSVF-multi-4的预测结构分别与三种已知表位特异性抗体进行对齐,分别对应II位点(PDB 3IXT)、IV位点(PDB 3O41)和V位点(PDB 5TPN)。c,筛选得到的多基序支架候选体的预测三维结构。d,展示各支架蛋白在起始温度和终止孵育温度条件下的圆二色谱谱图,用以评估其二级结构稳定性。e–g,采用表面等离子体共振技术测量四种支架蛋白分别与三种表位特异性抗体RSV90(V位点)、motavizumab(II位点)和101F(IV位点)之间的归一化稳态结合亲和力。
4 多表位支架在体内诱导位点特异性的中和抗体
在体外实验成功验证多表位设计之后,该研究进一步评估了这些多表位免疫原在体内是否能够诱导位点特异性的免疫应答。该研究假设,同时展示多种不同病毒表位的免疫原有望带来更高的免疫原性。此前为重构RSVF抗原表面,通常采用鸡尾酒疫苗策略,将三个中和位点分别支架化并呈现在三种不同的蛋白结构上。在这些设计中,真正与表位相关的残基仅占每种免疫原总表面积的20%至40%,这很可能导致相当比例的抗体反应指向宿主支架蛋白本身。相比之下,多表位设计将三个表位集中呈现在单一支架上,使表位相关表面积超过免疫原总表面积的50%,从而在减少支架“诱饵”表面的同时,理论上能够诱导更优的免疫应答。
为在体内评估免疫反应,该研究对初免小鼠分别使用三种支架进行免疫,包括单表位免疫原RSVFV-1以及两种多表位免疫原RSVF-multi-1和RSVF-multi-3。所有支架均被组装至自组装铁蛋白纳米颗粒上,以增强免疫原性。每组包含5只小鼠,采用同源加强免疫方案,连续注射三次RSVF前融合态三聚体或从头设计的纳米颗粒免疫原 (图4a)。另设异源免疫方案,先以RSVF进行初免,随后以PBS或纳米颗粒免疫原进行两次加强免疫 (图4a)。接受RSVF免疫的小鼠产生的抗体滴度和中和水平与既往研究结果一致 (图4b,f)。与此前关于单表位支架免疫原的研究相符,RSVFV-1主要诱导了针对自身结构的抗体,而对前融合态RSVF的交叉反应滴度较低,且未检测到明显的中和活性,表明诱导的抗体中有较大比例指向从头设计的支架本身 (图4b,c)。相比之下,两种多表位免疫原均诱导了中等水平的前融合态RSVF抗体滴度,较单表位免疫原提高了数个数量级 (图4c)。进一步利用单一免疫原探针检测表位特异性反应时,在两种多表位纳米颗粒免疫中均检测到可量化的位点特异性抗体滴度 (图4d)。尽管II位点的滴度未高于野生型RSVF免疫,但IV位点和V位点的抗体滴度显著提升。尽管针对支架表面或新生表位的抗体在一定程度上限制了整体RSVF抗体滴度,这些结果仍表明,多表位支架能够诱导针对三种被支架化表位的特异性抗体反应。此外,在同源免疫条件下,RSVF-multi-1免疫血清表现出一定的中和活性,尽管仍低于前融合态RSVF,但这一趋势与既往鸡尾酒疫苗研究结果一致 (图4e)。
在多表位加强免疫条件下,小鼠对RSVF的抗体滴度与未加强免疫组相当 (图4b);异源加强免疫小鼠对支架的反应水平也与同源免疫方案相近 (图4c)。然而,异源免疫方案下的位点特异性抗体滴度低于同源免疫方案 (图4f),凸显了在初免—加强免疫实验中使用异源支架时,定向增强特定表位反应所面临的困难。
综合包括体内实验在内的全部数据,该研究证实深度学习方法能够生成相较单表位设计具有更大免疫有效表面积的蛋白免疫原,并显著改善免疫原性及生理相关的抗体应答。这些结果也从实验层面验证了一个关键观点,即免疫原中表位所占相对表面积是决定免疫原性的重要因素。
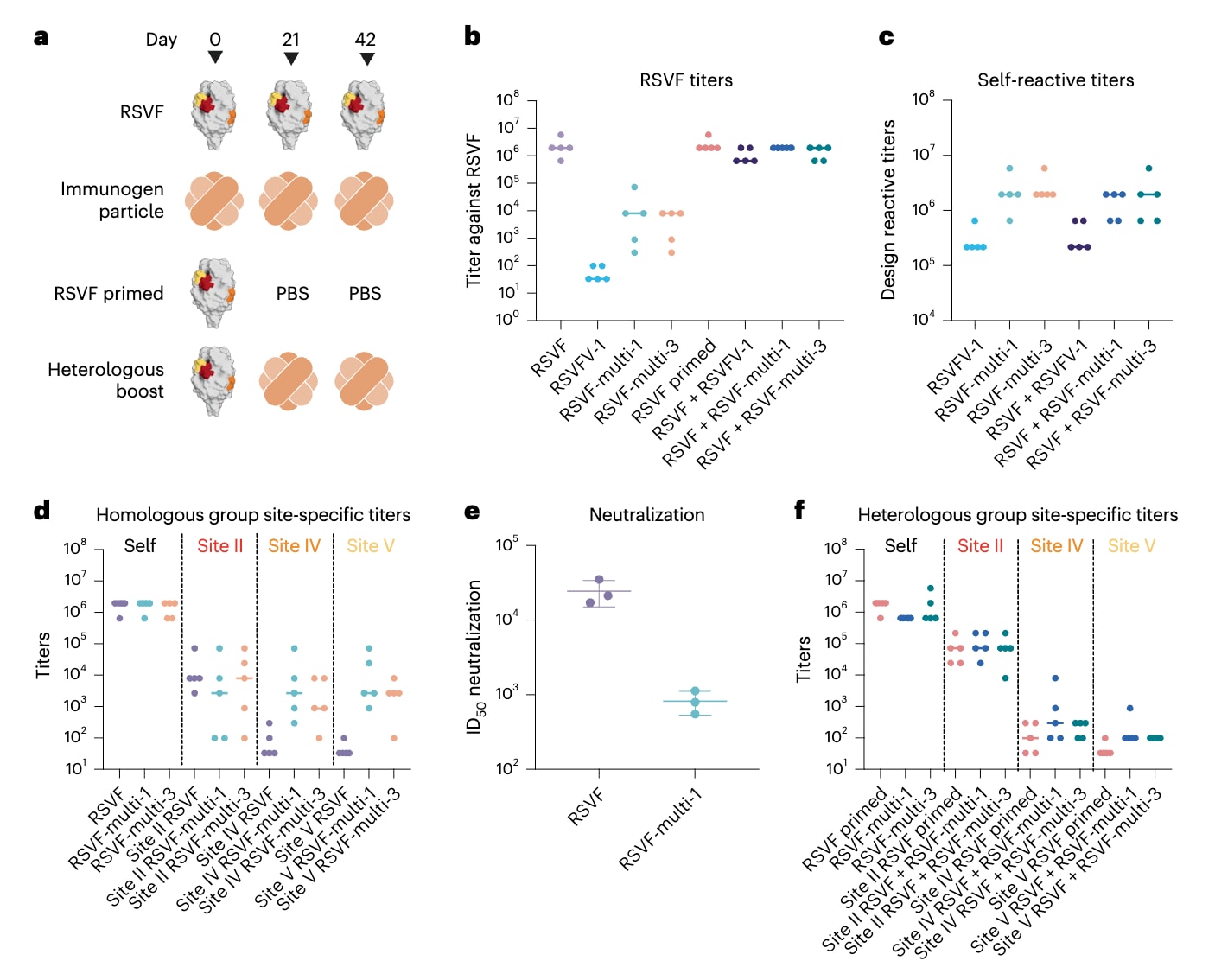
图4|单表位与多表位免疫原的免疫原性评估。 a,将小鼠分为四组(每组
5 多基序原位生成设计具有较高的结构精度
为评估多基序设计的结构准确性,该研究解析了两种RSVF多表位免疫原的晶体结构,分别为与motavizumab Fab形成复合物的RSVF-multi-1以及游离状态下的RSVF-multi-4,其分辨率分别为
在RSVF-multi-4的游离晶体结构中,所有被支架化表位与天然表位结构均表现出高度相似性,其主链均方根偏差均小于
总体而言,在所解析的X射线晶体结构中,所有被支架化表位均表现出高度逼真的结构复现,在多种情况下,其主链和
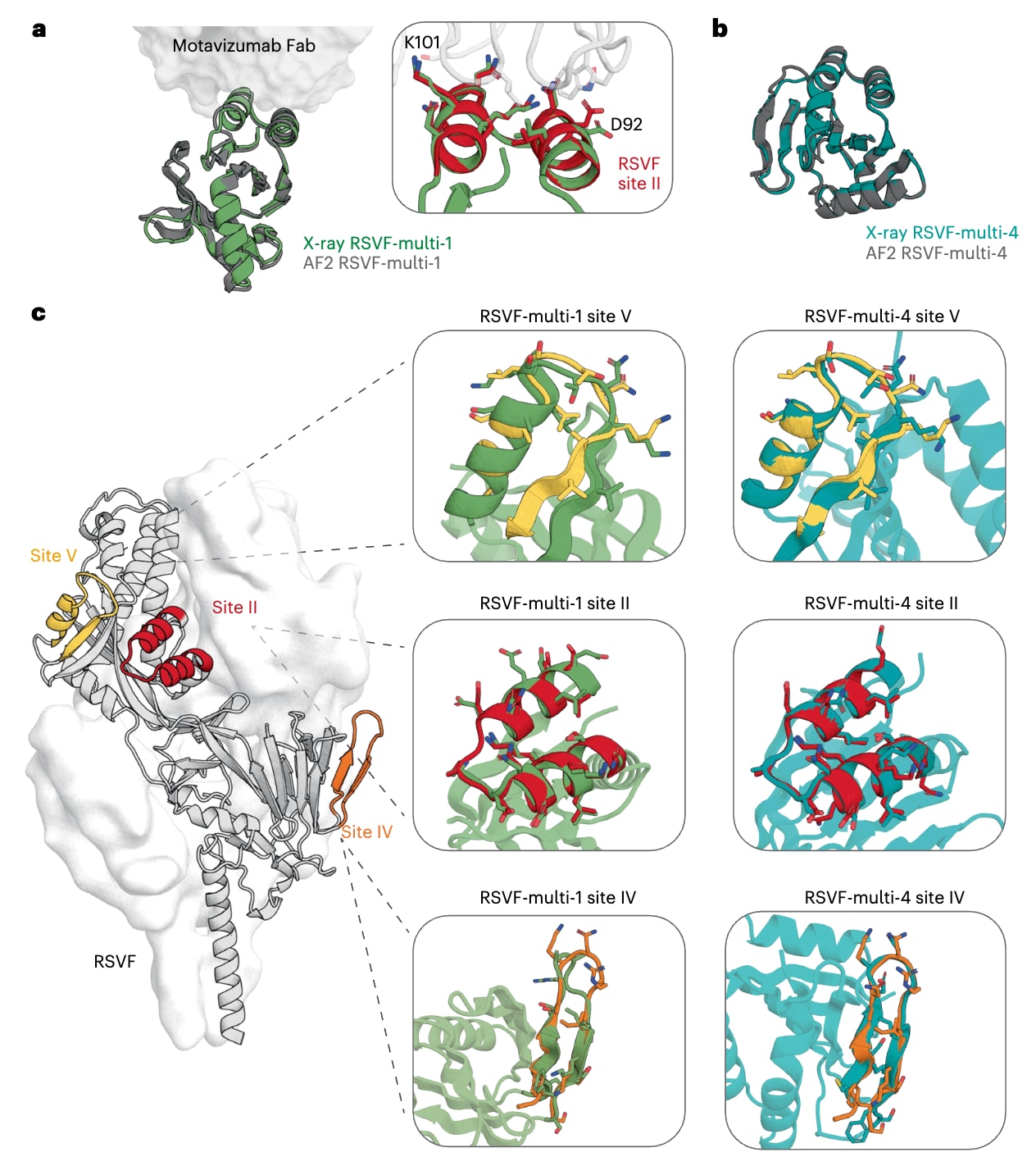
图5|单表位与多表位免疫原的结构表征。 a,RSVF-multi-1与motavizumab结合态的晶体结构(绿色)与AlphaFold2模型(灰色)的叠合结果。插图为局部放大视图,展示RSVF-multi-1中的主要接触残基与motavizumab结合态的天然RSVF II位点结构(红色,PDB 3O45)之间的叠合关系。b,RSVF-multi-4的晶体结构(青绿色)与AlphaFold2模型(灰色)的叠合结果。c,将晶体结构中解析得到的各表位分别与天然RSVF的V位点(黄色)、II位点(红色)和IV位点(橙色)结构进行叠合比较(PDB 5TPN)。
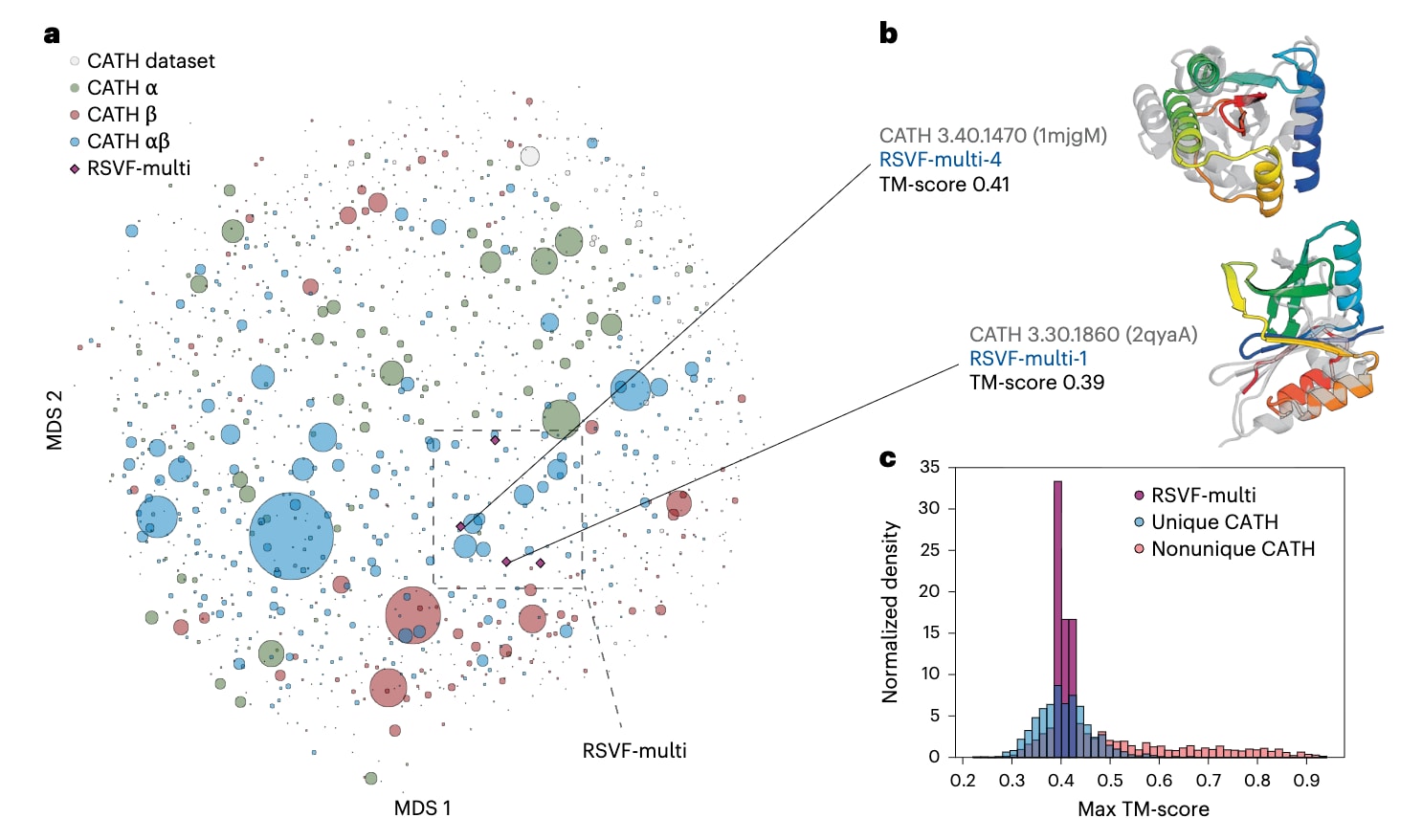
图6|呈现单表位与多表位的支架蛋白在结构上的独特性。 a,基于成对TM-score的多维尺度分析图,展示CATH拓扑家族代表结构与RSVF-multi设计体之间的结构关系,并按结构类型进行着色。圆点表示CATH中的不同拓扑类型,其相对大小反映该拓扑家族所包含的结构数量,紫色菱形表示四种多表位设计体。b,将经结构验证的多表位设计体RSVF-multi-1和RSVF-multi-4与其在CATH中最接近的拓扑家族代表结构进行对齐比较。c,将生成的独特拓扑与非独特拓扑在CATH拓扑家族中的最大TM-score分布进行比较,RSVF-multi设计体与其最接近结构同源体之间的TM-score落在独特拓扑的分布范围内,同时展示了RSVF-multi-1和RSVF-multi-4与最近CATH拓扑家族代表结构的对齐结果。
6 讨论
复杂结构基序的支架化设计长期受到可用于设计的结构空间限制,这使得从头蛋白设计在多数情况下只能针对单一设计问题给出单一基序的解决方案。与此不同,天然蛋白往往能够通过单条多肽链执行多种功能。该研究利用深度学习方法,实现了在非天然取向下将多达三种结构上彼此不同的基序嵌入同一从头设计支架中的目标。基于深度学习的基序支架化策略在结构与序列层面提供了更大的灵活性,使其能够容纳复杂表位,同时在寻找兼容支架时所需的人工干预显著减少,从而大幅提升了从头基序支架化设计的可及性。不同于以往依赖大规模文库甚至体外进化的单基序设计工作,该研究仅通过数量有限的设计序列,便获得了能够同时与三种抗体结合的多种设计体,这是迈向多功能蛋白设计的重要一步。通过在更大尺度上探索不同拓扑和序列,设计体的结合亲和力仍有进一步提升的空间。此外,该研究表明RFjoint2 Inpainting能够在保持高结构精度的同时,为多基序支架化问题生成具有不同基序相对取向的多样化拓扑解。所有三种表位均在与蛋白质数据库中已知结构显著不同的新型折叠中,实现了与天然表位在局部结构和功能上的高度一致。
作为免疫原,同时呈现多个抗原位点能够显著改善抗原表面的有效展示。所设计的多表位免疫原相比V位点单表位免疫原,诱导出更高水平的RSVF交叉反应抗体滴度,并且相较RSVF免疫血清表现出更优的位点特异性反应。异源加强免疫方案中,使用单表位支架作为探针并未观察到位点特异性滴度的进一步提升。一种潜在的替代策略是采用多表位支架进行初免,并使用包含相同嫁接表位但支架不同的多表位免疫原进行加强免疫,从而选择性增强针对目标表位的抗体反应,同时减少针对新生表位的抗体产生。此外,其中一种三表位免疫原表现出具有生理相关性的中和抗体滴度,说明与单表位免疫原相比,多表位免疫原能够通过单一成分介导针对更大抗原表面的免疫应答。既往研究已表明,采用单表位免疫原组成的鸡尾酒疫苗能够提升RSVF抗体滴度。相比之下,该研究提出的多表位免疫原为以单一组分替代鸡尾酒疫苗提供了一种可行方案,有望显著降低成本,并加快生产与验证过程。这类多表位设计相较单表位设计具有更高比例的理想抗原表面,能够在紧凑的支架上更真实地复现RSVF的抗原特征,同时降低诱导非目标抗体反应的风险。
除疫苗设计之外,将多个功能位点整合至单一蛋白中的能力在酶、传感器以及治疗性候选分子的设计中同样具有广泛应用前景。这些设计在结构上的高度新颖性以及与晶体结构之间的高度一致性,或许是迄今为止生成式深度学习方法在高度受限设计问题中构建精确定制化解决方案能力的最有力例证。