Nat. Rev. Immunol. 2011 | 对MHCⅠ类和MHCⅡ类抗原呈递的系统性理解

Neefjes, J.; Jongsma, M. L. M.; Paul, P.; Bakke, O. Towards a Systems Understanding of MHC Class I and MHC Class II Antigen Presentation. Nature Reviews Immunology 2011, 11 (12), 823–836. https://doi.org/10.1038/nri3084.
0 摘要
近三十年来,MHCⅠ类和MHCⅡ类分子介导的抗原加工与呈递的分子细节已得到广泛研究。尽管这些过程的基本原理大约在10年前已经得到阐明,但近年来的研究进一步揭示了大量细节,并为其调控机制和特异性提供了新的认识。MHC分子通过多种生物化学反应,将抗原片段有效呈递给免疫系统。这里对抗原呈递生物学的当前认识进行了及时评述,并概述了这一领域中仍被认为尚未解决的问题。随着越来越多的新发现不断纳入对MHCⅠ类和MHCⅡ类抗原呈递生物学的理解,这一体系逐渐展现为一个涉及多种细胞生物学过程的系统,本综述对此进行了讨论。
1 引言
MHCⅠ类和MHCⅡ类分子在功能上相似:它们分别在细胞表面向CD8+T细胞和CD4+T细胞呈递肽段。这些肽段来源不同——MHCⅠ类分子主要呈递细胞内来源的抗原,而MHCⅡ类分子主要呈递外源性抗原——并且通过不同的途径产生。两条途径之间还存在一种有趣的联系,即所谓的交叉呈递,在这一过程中,外源性抗原可由MHCⅠ类分子呈递。此外,当胞质内源性蛋白通过自噬或其他途径被降解时,这些蛋白也能够由MHCⅡ类分子呈递。病原体为操纵MHCⅠ类和MHCⅡ类通路而进化出的多种机制,也为理解抗原呈递生物学提供了新的线索;不过,这些内容近期已有综述,这里不再展开讨论。本文首先讨论MHCⅠ类抗原呈递,随后介绍MHCⅡ类抗原呈递。行文先概述基本通路,再结合该领域的最新进展,逐步形成对MHC分子抗原呈递的系统性描述。此外,还邀请了抗原加工与呈递领域的研究者,就若干尚未解决的问题提出看法。
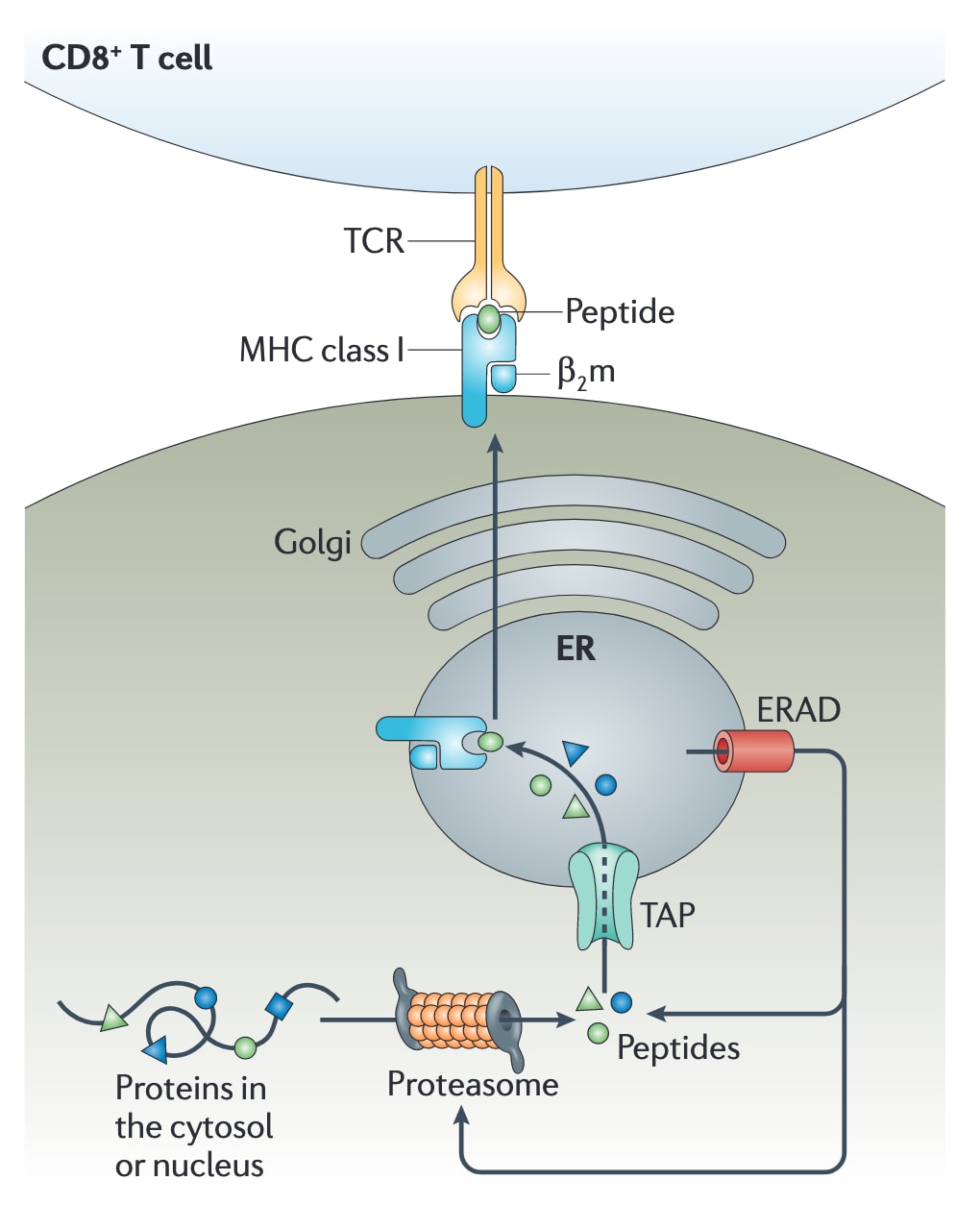
图1|MHCⅠ类抗原呈递的基本通路。 MHCⅠ类分子对细胞内抗原肽的呈递,是由一系列连续反应共同完成的。首先,抗原在蛋白酶体中被降解。随后,产生的肽段通过抗原加工相关转运体TAP被转运至内质网(ER)腔内,并装载到MHCⅠ类分子上。形成的肽-MHCⅠ类复合物随后从内质网释放,经高尔基体运输至质膜,最终在细胞表面呈递给CD8+T细胞。β2m,β2-微球蛋白;ERAD,内质网相关蛋白降解;TCR,T细胞受体。
2 MHCⅠ类抗原呈递的基本过程
MHCⅠ类分子表达于所有有核细胞表面,负责呈递来源于胞质和细胞核蛋白的片段。大量研究已经表明,抗原首先由胞质和细胞核中的蛋白酶体降解。产生的肽段随后在抗原加工相关转运体TAP的作用下被转运进入内质网,从而接触MHCⅠ类分子。在内质网中,MHCⅠ类异源二聚体由具有多态性的重链和一条称为β2微球蛋白(
在没有肽段结合时,MHCⅠ类分子的稳定依赖于内质网伴侣蛋白,如钙网蛋白、ERp57(又称PDIA3)、蛋白二硫键异构酶(PDI)以及专职伴侣蛋白tapasin。Tapasin能够与TAP相互作用,从而将肽段向内质网的转运过程与其递送至MHCⅠ类分子的过程耦联起来。当肽段与MHCⅠ类分子结合后,这些伴侣蛋白便会释放,完成组装的肽-MHCⅠ类复合物随后离开内质网,被运送到细胞表面进行呈递。相反,那些在内质网中未能彼此结合的肽段和MHCⅠ类分子,则会被送回胞质中降解。
3 MHCⅠ类通路的复杂性
近期的研究进一步凸显了这一基本通路中多个步骤的复杂性,具体如下。
3.1 抗原加工
MHCⅠ类分子所呈递的抗原,通常来源于那些已接近功能寿命终点、并经蛋白酶体介导降解的蛋白质。蛋白质的半衰期差异很大,从几分钟到数天不等。因此,按常理推测,某种蛋白从合成到其肽段被MHCⅠ类分子呈递出来的时间,应当与该蛋白本身的半衰期大致相当。然而,这一设想与一个重要现象并不一致:病毒抗原被MHCⅠ类分子呈递的速度,远快于其天然半衰期所允许的时间。
这一矛盾后来得到了解释。研究发现,相当大比例的新生蛋白——约占全部新合成蛋白的30%至70%——会在形成具有功能的成熟蛋白之前,于合成后立即被降解。这些蛋白被称为DRiPs,即核糖体缺陷产物。它们可能源于转录或翻译错误、可变阅读框的使用、未能成功组装进入更大的蛋白复合体、由于氨酰tRNA合成酶出错而掺入错误氨基酸,或者泛素修饰发生异常等情况。细胞会迅速降解这些DRiPs,以防止蛋白聚集体形成并损害细胞活性。正因为存在这种与翻译紧密耦联的降解途径,病毒蛋白的翻译也就直接与MHCⅠ类抗原呈递过程联系起来。DRiPs因此解释了为何流感病毒感染后约1.5小时,T细胞便可识别感染细胞,而不是等到约8小时后,待首批稳定病毒蛋白降解时才发生识别。
对MHCⅠ类抗原呈递的研究还揭示了另一个出人意料的现象,即某些被呈递的肽段并非直接由基因组编码。蛋白酶通常通过切割产生蛋白片段,但在某些情况下,它们也能够将两个肽段重新连接起来,例如蛋白酶体20S核心介导的蛋白剪接。由此产生的连接肽无法单纯依据基因组序列进行预测,其中还包括可被肿瘤特异性细胞毒性T淋巴细胞识别的新抗原。
脉冲-追踪实验表明,相当一部分MHCⅠ类分子始终无法结合合适的肽段,这些分子最终会通过内质网相关蛋白降解系统ERAD被清除。因此,肽段的浓度——而这又与蛋白质合成速率密切相关——实际上成为MHCⅠ类抗原呈递反应中的限速因素。蛋白质合成会受到多种刺激影响,例如干扰素γ、通过哺乳动物雷帕霉素靶蛋白
总体来看,MHCⅠ类分子所呈递的肽组中,有相当大一部分来自DRiPs,也就是那些从未真正具备功能的蛋白产物,而DRiPs的水平又会受到影响翻译过程的外界因素调控。此外,翻译机制本身的不完美,再加上肽段连接反应,还能够产生基因组中并不存在编码信息、却依然具有免疫原性的全新肽片段。因此,针对MHCⅠ类相关肽组的研究不仅拓展了对肽库组成的认识,也揭示了此前并未被充分认识的细胞生物学过程。
3.2 蛋白酶体
26S蛋白酶体由一个具有蛋白酶活性的20S核心桶状结构和两个19S帽状复合体组成。它产生了供MHCⅠ类分子呈递的大部分肽段,并决定这些肽段的羧基末端。由此生成的肽段随后通常只需再经过氨基端修剪即可。
目前已知还存在两类“替代性”蛋白酶体:一类是免疫蛋白酶体,主要表达于多种免疫细胞中;另一类是胸腺特异性蛋白酶体,主要表达于胸腺上皮细胞中。在这些蛋白酶体中,免疫细胞特异性或胸腺特异性的蛋白水解亚基会被整合进20S核心桶状结构,从而改变蛋白酶体的降解模式。过去通常认为,免疫蛋白酶体能够通过选择性地产生某些特定的、具有免疫原性的肽段,从而提高MHCⅠ类抗原呈递效率。然而,蛋白酶体实际上必须为各种不同的MHCⅠ类等位基因产生肽段,而不同等位基因可结合的肽序列并不相同。因此,很难设想免疫蛋白酶体能够同时提升所有MHCⅠ类等位基因所需肽段的生成,因为一种肽段生成增加,往往意味着另一种肽段的生成会相应减少。
最近,研究揭示了免疫蛋白酶体一个出乎意料的功能。免疫应激以及IFNγ暴露会诱导活性氧
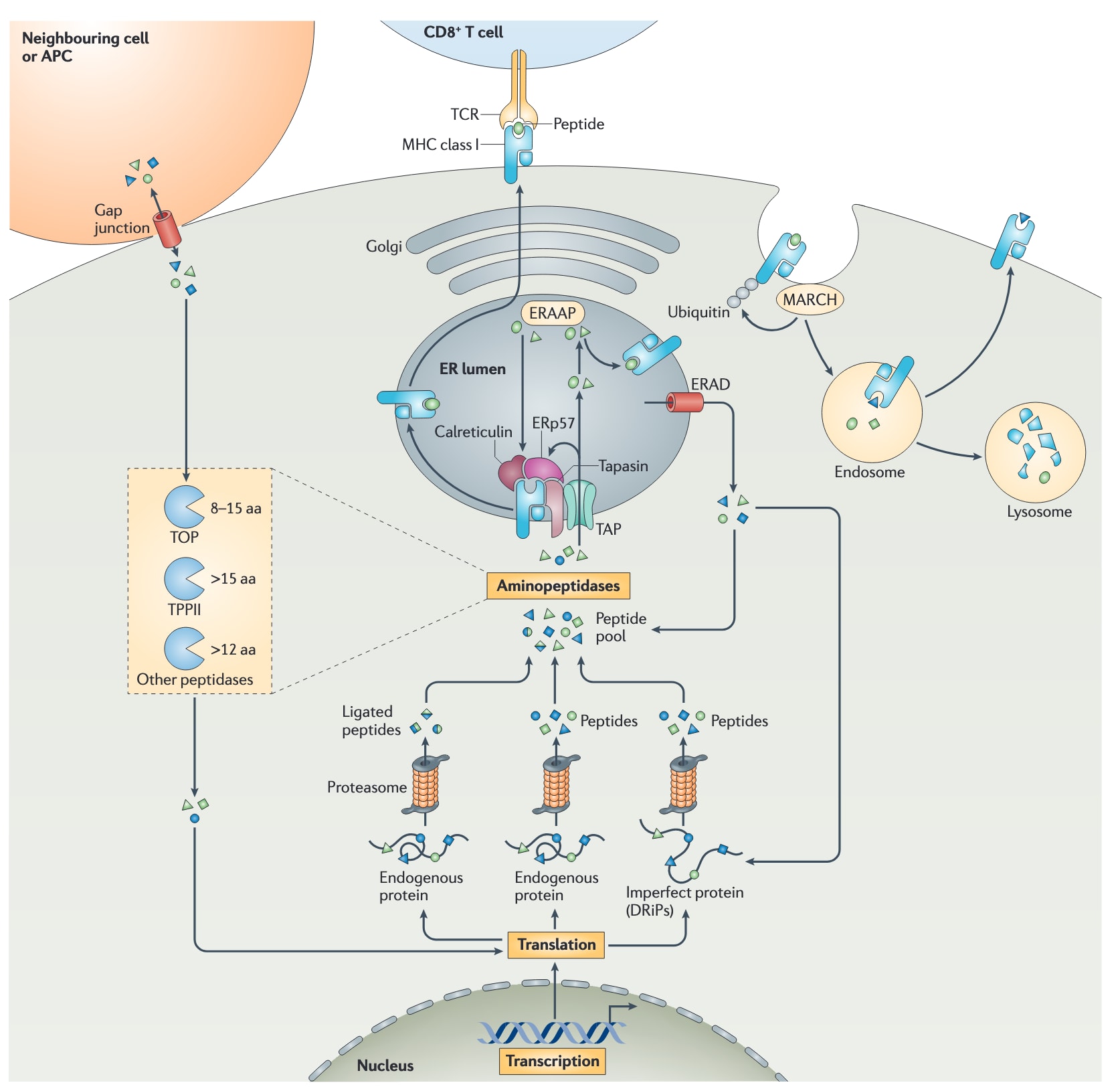
图2|MHCⅠ类抗原呈递通路的复杂性。 抗原的生命周期始于转录和翻译。许多蛋白质在合成过程中并不完全正确,这些产物被称为核糖体缺陷产物DRiPs。与具有功能的内源性蛋白类似,DRiPs也会在细胞核和胞质中被蛋白酶体降解。蛋白酶体还能够通过连接反应生成新的、并非由遗传编码直接决定的抗原。由此产生的肽段会成为胞质氨肽酶的底物,例如thimet oligopeptidase(TOP)和tripeptidyl peptidase II(TPPII),这些酶会对肽段进行修剪,并降解其中的大多数。只有一小部分肽段能够逃避末端降解,并通过抗原加工相关转运体TAP转运进入内质网(ER)腔。TAP是肽装载复合体PLC的一部分,PLC还包含MHCⅠ类分子、ERp57、钙网蛋白和tapasin。肽段可以直接与MHCⅠ类分子结合,也可能需要先经抗原加工相关内质网氨肽酶ERAAP进一步修剪,然后才会在PLC内或PLC外被认为适合与MHCⅠ类分子结合。形成的肽-MHCⅠ类复合物随后从PLC和内质网中释放,并被转运至质膜,以向CD8+T细胞呈递抗原。那些未能成功相遇的肽段和MHCⅠ类分子,会通过内质网相关蛋白降解系统ERAD被送回胞质。在胞质中,它们会被肽酶和蛋白酶体进一步修剪或降解。一部分肽段还可经由缝隙连接转移进入邻近细胞,从而进入这些细胞中的MHCⅠ类通路。细胞表面的MHCⅠ类分子可被MARCH家族蛋白泛素化,从而促进其内吞并进入溶酶体降解。另有一部分被内吞的MHCⅠ类分子,在与内体肽段发生交换后可被再循环利用。APC,抗原呈递细胞。
3.3 经修饰理解后的MHCⅠ类肽装载复合体
肽段一旦生成,就必须接触到那些在内质网中等待装载肽段的MHCⅠ类分子。肽转运体TAP负责将胞质中的肽段泵入内质网腔。由于TAP并不位于核膜的核侧表面,因此来源于细胞核的肽段必须先扩散到胞质中,随后才能遇到TAP。TAP同时还作为MHCⅠ类分子折叠的平台,因为它能够结合一个或多个tapasin分子。Tapasin是MHCⅠ类分子的专职伴侣蛋白。在内质网中,MHCⅠ类分子处于部分折叠状态,并在另外两种常见伴侣蛋白——钙网蛋白和ERp57——以及可能参与肽结合的伴侣蛋白PDI的帮助下得到稳定。钙网蛋白是一种凝集素,可结合发生N-糖基化的MHCⅠ类分子,从而增强MHCⅠ类分子与tapasin之间的相互作用稳定性。由TAP、tapasin、MHCⅠ类分子、ERp57和钙网蛋白组成的复合体,被称为肽装载复合体,即PLC。PLC能够确保肽段高效装载到MHCⅠ类分子上。当MHCⅠ类分子结合了亲和力足够高的肽段后,这一复合物便从伴侣蛋白上释放出来,通过内质网质量控制系统,最终表达于质膜表面。
Tapasin还可能具有肽编辑功能,其作用与MHCⅡ类呈递过程中HLA-DM所发挥的作用相似。研究显示,tapasin的存在会使装载到MHCⅠ类分子上的肽段种类更加多样,但这些肽段的平均亲和力反而较低。其背后的机制目前仍令人困惑。
TAP可将长度为8至16个氨基酸的肽段转运入内质网。这些肽段在与MHCⅠ类分子结合之前,可能还需要在内质网中进一步修剪。目前在小鼠中已鉴定出一种内质网氨肽酶,称为抗原加工相关内质网氨肽酶ERAAP;在人类中则发现了两种,分别称为ERAP1和ERAP2。由于ERAAP并不属于PLC的一部分,许多肽段必须先在PLC之外的内质网腔中被修剪,然后再重新进入PLC与MHCⅠ类分子结合,或者在PLC之外寻找游离的MHCⅠ类分子。ERAAP很可能将肽段修剪至最短8个氨基酸,这一长度仍适合装载到MHCⅠ类分子上。当这些肽段未能成功结合MHCⅠ类分子时,它们会通过ERAD途径被清除,重新进入胞质并被彻底降解,或者再次经历TAP转运和MHCⅠ类分子筛选的循环(图2)。
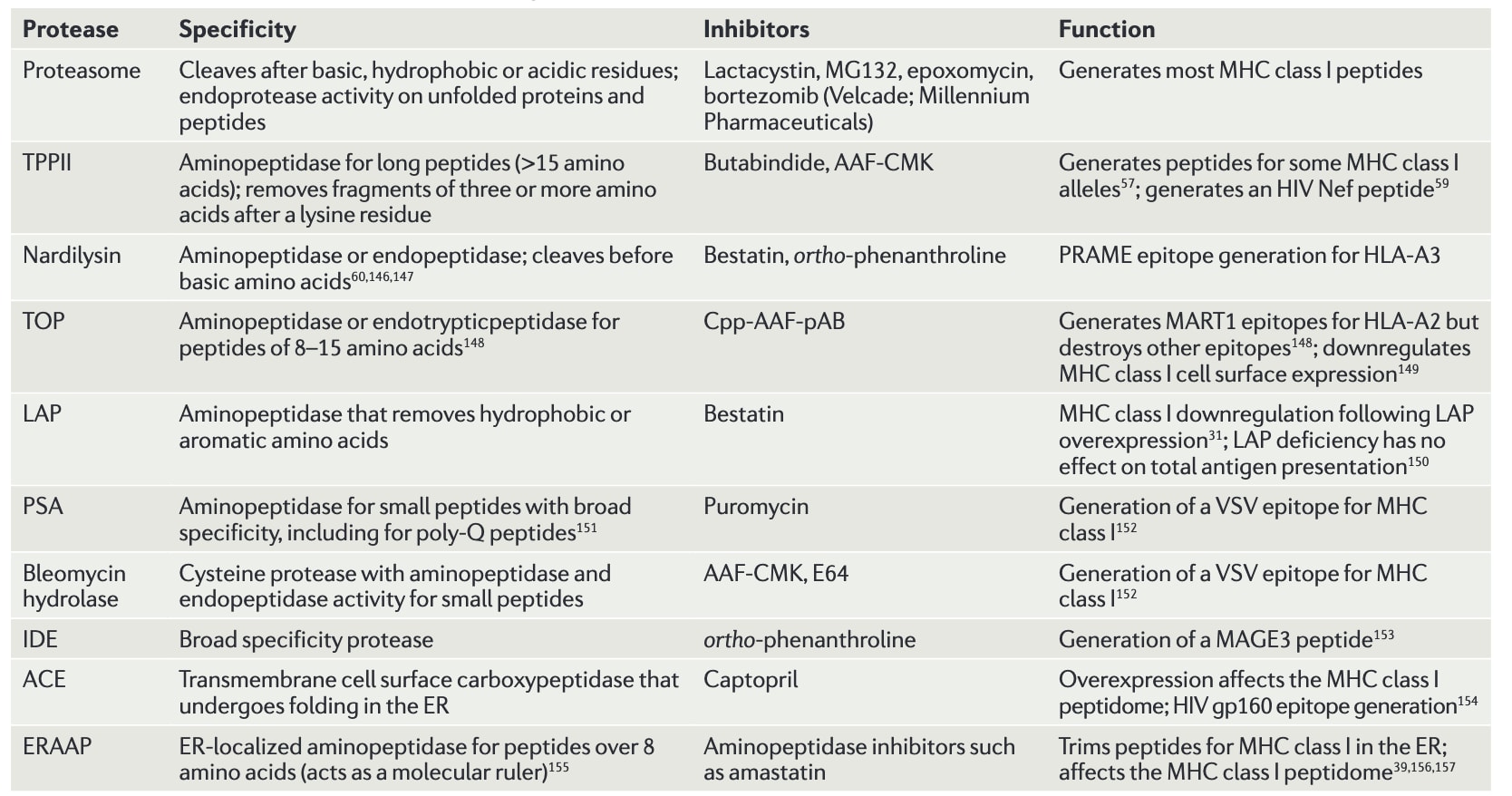
表1|MHCⅠ类通路中的蛋白酶
3.4 邻近细胞介导的抗原呈递
MHCⅠ类通路原本是为了呈递感染细胞内部产生的抗原肽而设计的。这样的安排有助于避免附带损伤,也就是减少对无辜邻近细胞的误杀。然而,许多细胞之间通过缝隙连接相互联通,这类连接允许小分子胞质成分或离子,如ATP、Ca2+以及肽段,进入邻近细胞的胞质中。这样一来,肽段就能够进入相邻细胞的抗原呈递通路。缝隙连接在细胞凋亡早期阶段仍然保持稳定,直到细胞膜起泡发生之前都是如此。因此,垂死细胞中由caspase产生的肽段,能够转移到尚未发生凋亡的邻近细胞中,并在那里完成抗原呈递。
在淋巴结中,树突状细胞DC通过与其他细胞接触可以形成缝隙连接,这可能促进交叉致敏。在小鼠中,缝隙连接还可使肿瘤细胞中的肽段转移到抗原呈递细胞APC中,从而诱导强烈的抗肿瘤免疫反应。因此,MHCⅠ类抗原呈递通路似乎并非完全封闭于单个细胞内部,而是能够与直接相邻的细胞共享抗原信息,只是这样做的代价,是在一定程度上增加了对无辜旁观细胞的误伤风险。
3 MHCⅠ类分子的多态性
在大多数物种中,MHCⅠ类重链由3个基因编码,在人类中分别是HLA-A、HLA-B和HLA-C。这3个位点都具有高度多态性,这也是MHC分子最鲜明的特征。MHC蛋白的多态性会导致肽结合槽结构不同,由于肽段对接所依赖的锚定位点残基存在差异,不同的MHCⅠ类分子因而能够识别各自特定的肽段。围绕MHCⅠ类基因座及其等位基因的深入研究还揭示了更多层面的差异。通常,HLA-A和HLA-B的表达水平高于HLA-C。人类HLA-C以及小鼠中的H2-L之所以表达较低,涉及多个原因,包括微小RNA介导的转录和/或转录后调控,以及可结合的肽库较为受限,从而限制了其装配。
在人类中,3个位点产物之间的差异研究得最为充分。HLA-A和HLA-C能够高效结合PLC,而HLA-B分子则不能。然而,HLA-B的肽装载和向质膜的转运速度却快于HLA-A和HLA-C。这提示,与PLC结合并不一定会促进更快的肽装载,相反,HLA-A和HLA-C可能由于tapasin的编辑作用而对更多肽段进行取样,从而延迟肽-MHCⅠ类复合物从PLC中的释放。
MHCⅠ类基因座所编码的不同多态形式,其行为方式也并不相同。有些形式几乎总是能够成功装载肽段,例如人类中的多数HLA-B等位基因;而另一些变体中,只有一部分分子能够完成装载,对于HLA-A和HLA-C来说,这一比例约为30%至70%。一种可能的解释是,那些在正常条件下未能获得肽段的MHCⅠ类分子,实际上为病毒感染或应激状态下新产生的肽段预留了空间。如果确实如此,这类MHCⅠ类等位基因就可能更容易呈递免疫优势肽,这或许能够解释为什么某些HLA-A和HLA-C等位基因能够与HLA-B等位基因协同,共同增强对HIV感染的防护作用。
某些MHC等位基因与疾病之间的关联,其机制往往仍不清楚。例如,HLA-B27与强直性脊柱炎之间存在显著关联。对此已经提出了多种解释,包括HLA-B27重链二聚体在质膜上的表达,以及其所呈递肽段具有更高稳定性等。最近,一项涵盖大规模患者队列的研究还发现,HLA-B27阳性患者中ERAP1序列的变异也会促进强直性脊柱炎的发生。携带ERAP1突变的细胞在体外显示出约40%降低的肽酶活性,这提示针对HLA-B27的异常肽加工过程可能参与了这种自身免疫病。总体而言,尽管MHCⅠ类分子的抗原呈递路径在一般层面上已经较为清楚,但这些分子多态性所带来的后果,目前才刚刚开始被真正认识,而这些差异很可能正是解释感染易感性和自身免疫疾病关联性的关键。
4 MHCⅠ类通路中的数量关系
对于任何一个功能性反应而言,底物、酶和产物之间都必须保持平衡。对MHCⅠ类呈递来说,这一问题本质上与数量有关。一个细胞大约含有
这一现象之所以可能,是因为肽段在活细胞胞质中的半衰期仅有6到10秒。这意味着绝大多数肽段在接触TAP之前就已经被降解,从而避免了呈递队列的形成。胞质中的肽段会被多种氨肽酶降解。三肽基肽酶II即TPPII以及neurolysin能够切割较大的肽片段。较小的8至15个氨基酸长度的肽段则主要由TOP处理,更短的肽段则由其他几种肽酶继续分解。缺失这些肽酶的小鼠通常并不会表现出整体抗原呈递的明显异常。这可能是因为不同肽酶之间能够相互补偿,而不够精确的N端修剪也总可以由内质网氨肽酶ERAAP进一步校正。不过,尽管这些胞质肽酶对整体肽库并非必需,它们对于某些特定MHCⅠ类肽段的生成却是不可缺少的。
如果将这些肽酶强大的降解能力纳入MHCⅠ类抗原呈递的反应框架中,就会得到一些很有意思的推论。例如,为了说明为什么蛋白酶体产生的输入肽数量在PLC处成为限制因素,必须假定最终逃脱降解并成功与MHCⅠ类分子结合的胞质肽比例低于0.1%。这意味着,拷贝数低于1000的蛋白通常不会被免疫系统识别,因为除非其肽段异常稳定,否则很难在多种肽酶的联合作用下存活下来。
若要选择性提升病原体来源肽段的呈递效率,一种办法可能是将蛋白生成与降解过程耦联,并使其高效进入MHCⅠ类通路。例如,蛋白酶体可能定位在靠近TAP的位置,从而将肽段直接递送给TAP,尽量减少肽酶池的破坏作用,虽然目前尚无直接证据支持这一设想。此外,抗原也可能由某些偏向于促进呈递的“免疫核糖体”产生,但这些免疫核糖体究竟如何区分会生成抗原的蛋白和普通蛋白,目前仍不清楚。最近一项研究鉴定出一种核糖体相关伴侣蛋白BAG6,它能够在翻译过程中追踪错误定位的内质网蛋白,并将这一过程与蛋白酶体降解相连接。完全可以设想,还存在更多将翻译与降解耦联起来的系统,未来这些机制的揭示将有助于解释两者如何相互联系,以及如何生成DRiPs以支持快速抗原呈递。
当前逐渐形成的图景,更接近一个随机模型:系统输入的是大量过剩的肽段,最终输出的只是极少数幸存下来并结合于MHCⅠ类分子上的肽段。肽段生成的位置、细胞内全部肽酶池的作用,以及可能存在的伴侣蛋白调节,究竟如何共同塑造适用于MHCⅠ类抗原呈递的肽池,目前仍不清楚。正是通路中的这一环节,使得被呈递肽段无法实现绝对准确的预测,因为现有预测方法仍主要依赖于MHCⅠ类锚定位点残基的信息,以及TAP和蛋白酶体的特异性。
表2|MHCⅠ类抗原呈递中的未解问题

5 MHCⅠ类分子生命周期的终点
由于肽编辑作用和内质网质量控制系统的存在,细胞表面的MHCⅠ类复合物通常相对稳定。然而,质膜上也存在游离的MHCⅠ类重链。它们应当来源于MHCⅠ类异源二聚体的解离,因为在缺乏
与完整复合物相比,游离的MHCⅠ类重链在细胞表面的半衰期更短,这提示细胞内可能存在一套专门机制,能够识别不同形式的MHCⅠ类分子并将其导向降解。对单纯疱疹病毒免疫逃逸机制的研究,为这种降解方式提供了一种可能解释。这类病毒编码两种跨膜蛋白K3和K5,其携带RING结构域,能够泛素化靶蛋白的胞质尾部,从而促进这些蛋白发生内吞并进入溶酶体降解。它们实际上模拟了人类MARCH蛋白家族的作用,而MARCH4和MARCH9已被证明能够调控MHCⅠ类分子的半衰期。至于这些蛋白究竟如何识别MHCⅠ类分子,或者识别其游离重链形式,目前仍不清楚。
当MHCⅠ类分子被内吞进入内体后,它们实际上进入了经典的MHCⅡ类呈递通路。随后,酸化环境会促进原先结合的肽段释放出来,这些肽段可以被内吞途径中新生成的肽段所替换。部分MHCⅠ类分子会像MHCⅡ类分子一样被再循环到细胞表面,从而呈递来源于内体途径的抗原片段。正常条件下,这一通路并不占主导地位,因为从MHCⅠ类分子中分离到的内体来源肽段数量很少;不过在交叉呈递条件下,这一途径可能会更加常见。
总体来看,MHCⅠ类抗原呈递过程已经被解析到极为细致的程度(图2)。近年来,关于被呈递肽段类型以及系统中各组成成分功能的大量发现,显著加深了对这一过程的理解。许多步骤的特异性已经得到阐明,其中最典型的是肽段与MHCⅠ类分子之间的结合特异性。然而,尽管目前对MHCⅠ类抗原呈递各个环节已经积累了丰富认识,并且能够判断哪些肽段会被排除在MHCⅠ类结合之外,却仍然无法准确预测究竟哪些肽段最终会被呈递。免疫优势肽是如何产生的,它们是否以及如何不同于普通肽段,目前也尚无定论。这表明,要想让对MHCⅠ类抗原呈递的研究从描述性生物化学真正走向预测性生物化学,仍需填补若干关键的认识空白。
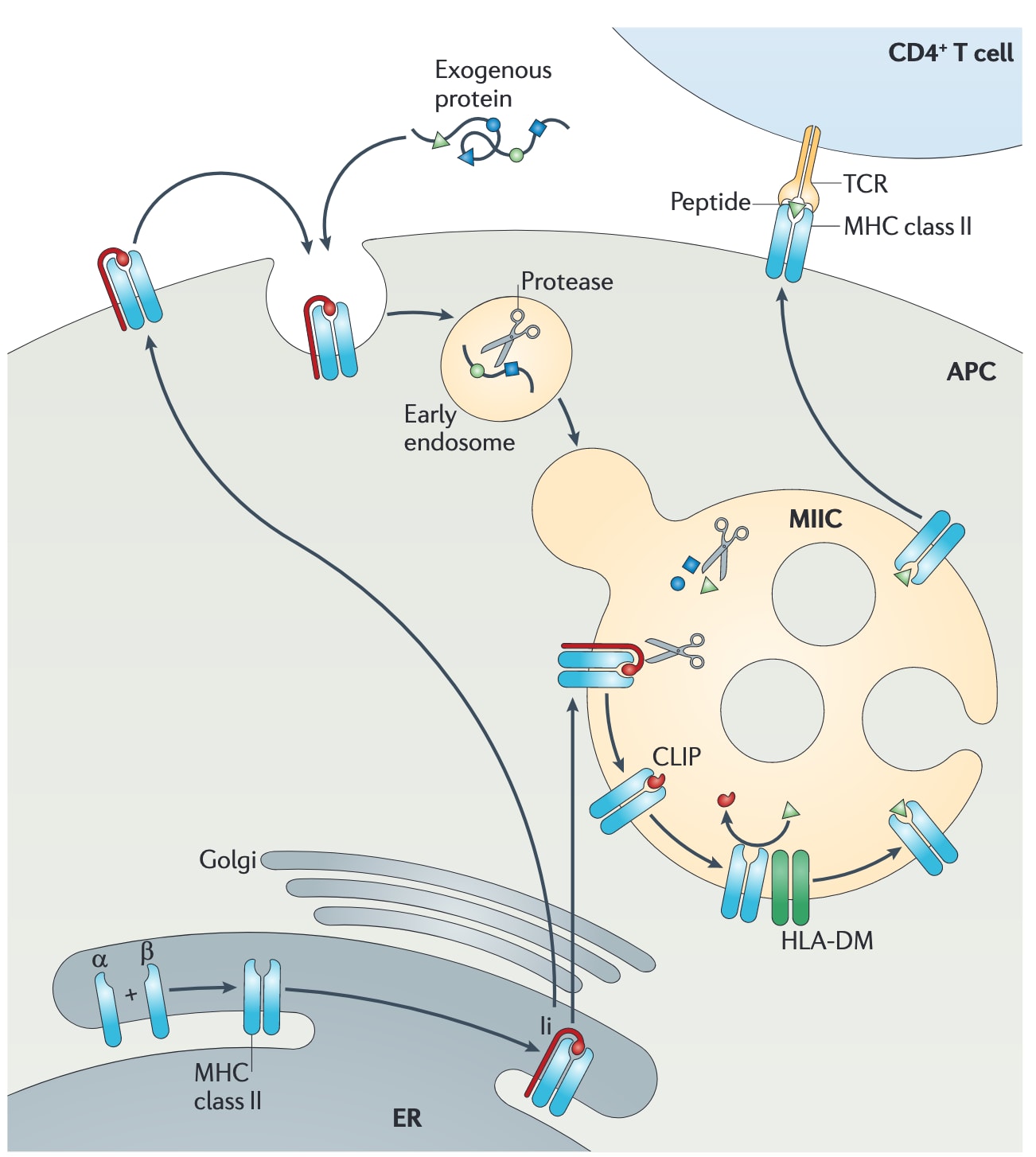
图3|MHCⅡ类抗原呈递的基本通路。 MHCⅡ类分子的
6 MHCⅡ类分子与MHCⅠ类分子的比较
之所以在同一篇综述中讨论MHCⅠ类和MHCⅡ类分子,是因为两者在许多特征上彼此重叠。二者都具有高度多态性,都拥有相似的三维结构——这与它们源自同一个祖先基因并通过简单基因复制产生有关——都定位于同一基因座区域,而且都承担着向免疫系统呈递肽段的相似功能。不过,这两类分子在组织分布上并不相同,所呈递的抗原肽类型也不同,原因在于它们依赖的是不同的细胞生物学通路。下面主要讨论MHCⅡ类抗原呈递通路与MHCⅠ类通路有何不同,以及这种差异如何使MHCⅡ类分子呈递来源于不同细胞区室的蛋白片段。
与MHCⅠ类重链类似,MHCⅡ类分子同样由3个多态性基因编码,在人类中分别是HLA-DR、HLA-DQ和HLA-DP,它们能够结合不同的肽段。许多MHCⅡ类等位基因是若干自身免疫病最强的遗传标志,这可能与它们所呈递的肽段有关。尽管不同MHCⅡ类变体与伴侣蛋白HLA-DM的相互作用方式似乎并不相同,但与MHCⅠ类相比,关于MHCⅡ类多态性如何影响其功能的研究仍然相对不足。下文所述的MHCⅡ类通路,主要建立在对HLA-DR以及小鼠MHCⅡ类分子
6.1 MHCⅡ类抗原呈递的基本过程
与广泛表达的MHCⅠ类分子不同,MHCⅡ类分子主要表达于专业抗原呈递细胞,例如树突状细胞、巨噬细胞和B细胞。大量研究已经表明,MHCⅡ类分子的跨膜
除去交叉呈递这一特殊情形,MHCⅠ类分子主要呈递来源于胞质的肽段,而MHCⅡ类分子则主要结合那些在内吞途径中降解蛋白所得的肽段。两者特异性的结合,几乎覆盖了来自所有细胞区室的抗原。不过,这些通路之间存在的关键差异,也使这一基本图景变得更加复杂。此外,这一领域仍有若干问题尚未厘清。目前还无法计算导致MHCⅡ类分子完成肽装载的各步反应效率,因为现有研究尚未提供所需的定量数据。
7 MHCⅡ类通路的复杂性
7.1 MHCⅡ类分子的表达
与MHCⅠ类分子的表达不同,MHCⅡ类分子的表达通常局限于抗原呈递细胞。不过,在非抗原呈递细胞中,MHCⅡ类分子的表达也可被IFNγ及其他刺激诱导,例如间充质基质细胞、成纤维细胞和内皮细胞中都可以出现这种诱导表达。在克罗恩病和嗜酸性食管炎等病理状态下,上皮细胞和肠神经胶质细胞也可表达MHCⅡ类分子。此外,某些皮肤病,如银屑病,还能诱导角质形成细胞表达MHCⅡ类分子。非抗原呈递细胞可在缺乏共刺激分子的条件下表达MHCⅡ类分子,从而维持外周耐受。那么,抗原呈递细胞与非抗原呈递细胞中的MHCⅡ类表达究竟是如何受到调控的呢?
MHCⅡ类表达的主调控因子是MHCⅡ类转录激活因子CIITA。CIITA通过MHCⅡ类增强体复合物被募集至MHCⅡ类基因座中的X1、X2和Y盒元件。该增强体复合物包含cAMP反应元件结合蛋白CREB、核转录因子Y即NFY以及调节因子X复合体RFX。相比之下,CIITA自身的表达调控要复杂得多,可产生I、III和IV等不同亚型,并分别在不同细胞类型中表达。
在树突状细胞中,MHCⅡ类基因座的转录控制还存在额外一层调节。在未成熟树突状细胞中,I型CIITA启动子可被4种因子结合,即PU.1、干扰素调节因子8即IRF8、核因子κB即NF-κB和SP1。这导致CIITA高水平转录,并进一步引起MHCⅡ类基因的高水平转录。随着树突状细胞成熟,这一复合体会被另一个含有PR结构域锌指蛋白1即PRDM1,又称BLIMP1的复合体所取代,后者抑制CIITA转录(图4b)。此外,CIITA在抗原呈递细胞中要发挥MHCⅡ类转录因子的功能,还需要先经历磷酸化和单泛素化修饰。
最近,研究者通过将全基因组小干扰RNA筛选结果与定量PCR分析结合,鉴定出CIITA的5个上游调控因子,分别是细胞分裂周期相关蛋白3即CDCA3、RMND5B、CCR4-NOT转录复合体亚基1即CNOT1、丝裂原活化蛋白激酶1即MAPK1以及含pleckstrin同源结构域家族A成员4即PLEKHA4。通过进一步分析这些因子彼此之间如何相互调节,研究揭示了一个控制CIITA及MHCⅡ类转录的复杂反馈机制(图4a)。事实上,只有这种复杂的转录反馈机制,才能解释一个主调控转录因子——在这里即CIITA——如何再被其后续受控因子层层反向调节。不过,构成这一反馈机制的各个因子本身也需要接受调控。进一步的系统生物学分析显示,CIITA表达的反馈调控是由转化生长因子
总体来看,MHCⅡ类基因座的转录由主调控因子CIITA控制,而CIITA又受到翻译后修饰以及多种主要但并非仅限于免疫细胞中活跃因子的调节。在特定信号环境和染色质修饰状态下,CIITA和MHCⅡ类分子也可以在非免疫细胞中表达,这种情况通常发生于感染或炎症过程中。
7.2 MHCⅡ类分子从内质网到MIIC的运输
尽管MHCⅠ类和MHCⅡ类分子都在内质网中组装,MHCⅠ类分子必须装载肽段后才能离开内质网,而MHCⅡ类分子则是通过与Ii结合而离开内质网。Ii存在4种不同剪接变体,差异表现为胞质尾部不同的p33和p35变体,或包含一个编码胱抑素家族蛋白酶抑制剂额外外显子的p43和p45变体。单独存在的MHCⅡ类
Ii的胞质尾部含有两个经典的双亮氨酸分选基序,它们负责将MHCⅡ类分子导向内体区室(图4c)。这些分选基序可被分选衔接蛋白AP1和AP2识别,其中AP1定位于反式高尔基网络,AP2则定位于质膜。因此,Ii既可以引导MHCⅡ类分子从反式高尔基网络直接运输至MIIC,也可以先到达质膜后再通过内吞进入MIIC。在人宫颈癌HeLa细胞和未成熟树突状细胞中,更偏好发生依赖AP2的内吞途径,而在成熟树突状细胞中,则可能以依赖AP1的直接分选为主。总体而言,Ii对于MHCⅡ类分子生命周期中的多个步骤都至关重要,不过其到达最终内体区室的路线可能并不唯一;而正是在这个终末区室中,Ii被降解,MHCⅡ类分子也终于获得了自己的肽段。
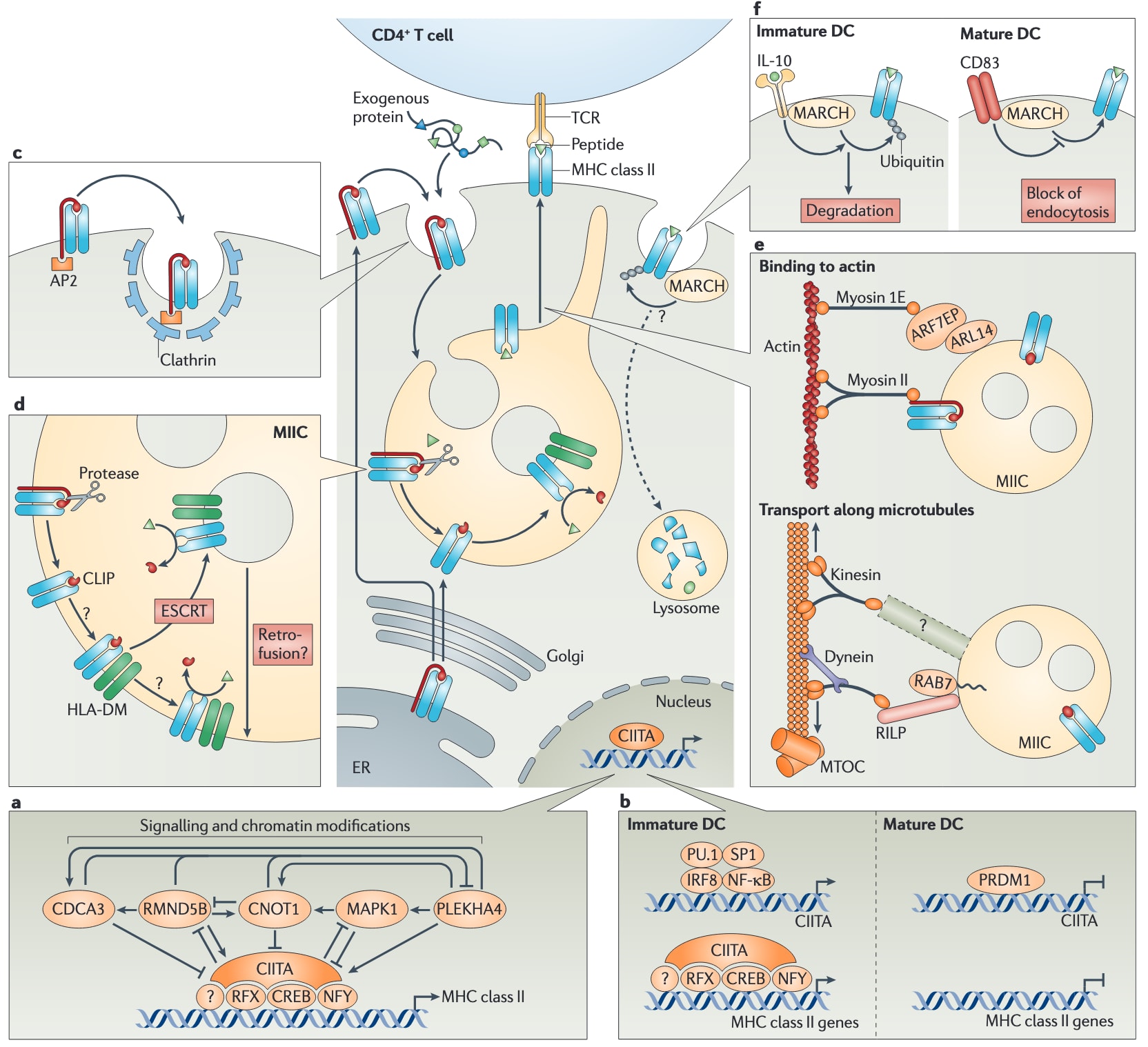
图4|MHCⅡ类抗原呈递通路的复杂性。 图中以多个模块展示了对MHCⅡ类通路各个步骤的认识,并将其投射到MHCⅡ类抗原呈递的基本通路之上。a|MHCⅡ类基因的转录受主调控因子MHCⅡ类转录激活因子CIITA控制,从而保证其组织特异性表达。CIITA的表达又受到一个反馈调控回路的影响,而这一回路进一步受两类普遍过程调节,即信号传导和染色质修饰。b|在未成熟树突状细胞中,CIITA的表达由一个激活性转录复合体调控,其中包含PU.1、SP1、IRF8和NF-κB;而在成熟树突状细胞中,CIITA则受到PR结构域锌指蛋白1(PRDM1)的抑制。因此,CIITA会与其他因子一起在未成熟树突状细胞中诱导MHCⅡ类基因转录,但在成熟树突状细胞中则不会。c|衔接蛋白AP2驱动Ii-MHCⅡ类复合物在质膜处进入网格蛋白包被小泡,并被运送至MHCⅡ类区室MIIC。d|在MIIC中,Ii被降解,MHCⅡ类分子随后与HLA-DM相互作用。大多数HLA-DM和MHCⅡ类分子定位并相互作用于由胞质侧内体分选复合体ESCRT machinery形成的内部结构中。位于内部的HLA-DM和MHCⅡ类分子是否会通过“逆向融合”返回到MIIC外膜,目前仍不清楚。e|MIIC或其管状延伸结构通过依赖微管的马达蛋白dynein和kinesin被运输至细胞膜。这些马达蛋白具有定位于MIIC的受体,例如作为dynein马达受体的RAB7相互作用溶酶体蛋白RILP。最终一步则涉及依赖肌动蛋白的肌球蛋白马达,它们可通过Ii与MIIC结合,因为Ii可与myosin II结合;也可通过GTP酶ADP-核糖基化因子样蛋白14(ARL14)与MIIC结合,后者会招募效应蛋白ARF7EP,而ARF7EP可作为马达蛋白myosin 1E的受体。后一种机制调控未成熟树突状细胞中的MIIC分泌。f|在未成熟树突状细胞中,MHCⅡ类分子自质膜的内吞可能需要泛素连接酶MARCH1参与,而MARCH1又受白细胞介素-10(IL-10)调控。成熟树突状细胞表面的CD83能够阻止MHCⅡ类分子的这种泛素化,从而稳定细胞表面的MHCⅡ类分子。CDCA3,细胞分裂周期相关蛋白3;CNOT1,CCR4-NOT转录复合体亚基1;CREB,cAMP反应元件结合蛋白;IRF8,干扰素调节因子8;MAPK1,丝裂原活化蛋白激酶1;NF-κB,核因子-κB;NFY,核转录因子Y;PLEKHA4,含pleckstrin同源结构域家族A成员4;RFX,调节因子X;TCR,T细胞受体。
7.3 MHCⅡ类分子的肽装载区室
MHCⅠ类分子是在内质网伴侣蛋白维持的部分折叠状态下与肽段结合的,而MHCⅡ类分子很可能并非如此,因为目前并不知道内体系统会参与蛋白折叠。自1990年通过电子显微镜首次观察到MIIC以来,MHCⅡ类分子究竟在何处完成肽装载,一直存在争议。当时研究显示,MIIC含有MHCⅡ类分子和Ii,具有多层板样形态,呈酸性,并含有溶酶体蛋白酶和CD63,因此被定义为一种晚期内体区室。此后又陆续鉴定出其他相关结构,这使得MIIC的定义需要不断修订。
随后发现,MHCⅡ类伴侣蛋白HLA-DM定位于晚期内体中,它能够稳定结合了CLIP或尚未结合CLIP的MHCⅡ类分子,从而在高亲和力肽段结合之前,防止MHCⅡ类分子聚集或被降解(图4d)。在这一发现基础上,进一步的研究又识别出晚期内体四跨膜蛋白、参与Ii降解的cathepsin S和cathepsin L等成分。体外重建实验显示,构成最小MIIC所需的分子仅包括MHCⅡ类、HLA-DM和组织蛋白酶。综合现有数据来看,只要某一晚期内体结构至少包含这三类因子,就可满足MIIC的基本标准。
使问题更复杂的是,MIIC并不是一个均质结构,而是表现出多种不同形态,包括多泡型、混合型和多层板型,它们可能代表了不同成熟阶段。MHCⅡ类、HLA-DM以及其他相关分子主要位于MIIC的内部结构中。要到达这些位置,它们必须先被泛素化,并通过内体分选复合体ESCRT machinery在MIIC外膜上完成分选。此外,荧光共振能量转移研究提示,HLA-DM与MHCⅡ类的相互作用主要发生在MIIC内部囊泡上,而不是在其外膜上。因此,目前一种常见设想是,携带MHCⅡ类分子和HLA-DM的内部囊泡会重新与MIIC外膜融合,从而使这些分子能够嵌入质膜,而不是以外泌体形式分泌出去。然而,这种“逆向融合”过程至今尚未在分子层面得到明确定义。
另一种模型则认为,树突状细胞表面出现的肽装载MHCⅡ类分子来源于MIIC外膜,而内部囊泡上的MHCⅡ类分子则注定要被降解。不过,由于大多数MHCⅡ类分子实际上都位于内部囊泡上,如果这些分子真的是降解命运,那么生物化学实验中理应观察到显著的MHCⅡ类损失,但实际并未发现这种现象。因此,是否存在逆向融合以及其分子机制究竟如何,仍需进一步解析,才能真正解决这一争议。
尽管目前普遍认为MHCⅡ类分子的肽装载主要发生在MIIC内,但仍有许多问题尚未解决。这些问题包括MHCⅡ类分子进入MIIC的具体机制,以及三聚体Ii在介导早期内体融合和调控MHCⅡ类胞内运输中的功能。MHCⅡ类分子如果在内吞通路不同区段完成装载,很可能会呈递不同的肽段,而HLA-DM在这些不同位置所发挥的帮助作用程度也可能并不一致。最后,在未成熟树突状细胞中,抗原降解会被延缓,这可能是一种使抗原得以长期储存并持续呈递的机制。也就是说,尽管最小MIIC的组成已经得到界定,但不同MIIC形态、不同蛋白水解活性、树突状细胞成熟过程中受控的酸化过程、逆向融合以及其他相关过程所带来的后果,仍需要进一步阐明,才能更完整地理解MHCⅡ类抗原装载这一胞内过程。
表3|MHCⅡ类抗原呈递中的未解问题

7.4 MHCⅡ类分子从MIIC到质膜的运输
像MIIC这样的晚期内体区室通常并不是典型的再循环结构,但MHCⅡ类分子、HLA-DM、四跨膜蛋白以及其他分子仍能够从MIIC被运输到质膜。MIIC中的内容物,包括MHCⅡ类分子在内,会在特定时间后从MIIC释放出来。这一释放过程受到多种因素调控,例如胆固醇、胞质
已知的一项机制是,MIIC沿微管由dynein驱动向内运输,这一过程受MIIC上的RAB7相互作用溶酶体蛋白RILP调控(图4e),而RILP又受到胆固醇感受器OSBP相关蛋白1L即ORP1L以及内质网驻留蛋白VAMP相关蛋白A即VAPA的控制。也就是说,内质网本身能够调控MIIC的运输,这也解释了胆固醇为何会影响MHCⅡ类抗原呈递。树突状细胞可能具有一定特殊性,因为在这类细胞中,含有MHCⅡ类分子的囊泡从MIIC向质膜的运输会受到成熟信号调控。这些信号会提高MHCⅡ类分子在细胞表面的表达水平,但代价是细胞内MHCⅡ类分子储库的减少。在树突状细胞中,脂多糖可诱导来源于MIIC的管状结构形成,从而构建出一个由运动中的囊泡和管道组成的复杂网络,这些结构可能都能够与质膜融合。
那么,树突状细胞中的MHCⅡ类运输究竟由什么控制?目前已有两类以肌动蛋白为基础的马达参与其中。其一是常见的肌球蛋白myosin II,它可能通过与Ii相互作用来调控树突状细胞中的MHCⅡ类运输(图4e)。另一条调控树突状细胞中MHCⅡ类运输的通路,则是通过整合siRNA筛选与细胞生物学分析发现的。研究首先筛选出影响MHCⅡ类表达的siRNA,随后鉴定出那些在树突状细胞成熟过程中表达下调的靶基因,最后再在未成熟树突状细胞中沉默这些剩余候选基因。结果发现,沉默其中某些基因后,MHCⅡ类分子的胞内分布会重新排列,呈现出与成熟树突状细胞相似的特征,尽管这些细胞在其他活化标志物方面仍保持未成熟状态。这些候选基因中包括编码GTP酶ADP-核糖基化因子样蛋白14即ARL14,又称ARF7的基因。ARL14定位于MIIC,并可招募效应蛋白ARF7EP,即先前称作C11ORF46的蛋白,后者可作为马达蛋白myosin 1E的受体。这一通路负责控制树突状细胞中MHCⅡ类分子的输出(图4e)。至于脂多糖诱导的成熟信号究竟如何调控这些通路,目前仍不清楚,但这种机制可能与其他溶酶体相关细胞器的诱导性分泌存在某种相似性,例如细胞毒颗粒、黑素体和Weibel-Palade小体。
7.5 MHCⅡ类分子的生命周期终点
与MHCⅠ类分子类似,MHCⅡ类分子也并非无限期存在。不过,MHCⅡ类复合物相对更加稳定,因为它们已经经历过晚期内体环境的筛选,并且不会在质膜上解离。此外,在树突状细胞成熟之后,MHCⅡ类分子的半衰期还会显著延长。那么,这些分子最终是如何被降解的呢?
MHCⅡ类分子与MHCⅠ类分子一样,也可被MARCH1泛素化。由于树突状细胞成熟时MARCH1的表达水平下降,同时MHCⅡ类分子的泛素化程度也随之降低,因此曾有观点认为,泛素化决定了MHCⅡ类分子的半衰期。白细胞介素-10可下调细胞表面MHCⅡ类分子的表达,并调控MARCH1的表达。此外,共刺激分子CD83在成熟树突状细胞中高表达,它可抑制MARCH1与MHCⅡ类分子之间的相互作用,从而阻止MHCⅡ类分子的泛素化。这些观察结果共同提示,泛素化与MHCⅡ类分子半衰期之间存在因果联系(图4d)。
不过,这一联系近来受到了挑战。研究构建了表达不能被泛素化的MHCⅡ类突变分子的小鼠,这些分子含有阻断泛素化的氨基酸替换。结果显示,这些小鼠的MHCⅡ类抗原呈递水平仍然基本正常,只是质膜上的MHCⅡ类表达略有升高。因此,也有可能MHCⅡ类分子的泛素化并不主要参与其内吞和降解,而是参与内吞通路中的另一过程,即内体途径内的分选。总体而言,MHCⅡ类分子具有极高的稳定性,但其半衰期仍具有细胞类型特异性。控制MHCⅡ类分子降解的机制尚未完全建立,其中可能涉及泛素化。最有可能的情况是,MHCⅡ类分子像其他溶酶体蛋白一样,最终通过溶酶体蛋白水解被降解,但细胞究竟依靠何种精确机制来区分“老”的MHCⅡ类分子和“新”的MHCⅡ类分子,目前仍未解决。
7.6 MHCⅡ类分子的outside-in信号传导
MHCⅡ类分子在向T细胞呈递肽段时,通常被看作介导inside-out信号,但近来的研究提示,MHCⅡ类分子本身也可作为信号受体发挥作用,从而介导outside-in信号传导。这样的信号可导致活化抗原呈递细胞发生凋亡,进而促使免疫反应终止。相反,在黑色素瘤细胞中,若MHCⅡ类分子与浸润淋巴细胞表面表达的配体LAG3结合,则可通过激活存活通路来阻止细胞死亡。
由于MHCⅡ类分子的胞质尾很短,而且并不含有可识别的信号基序,因此必须有衔接分子参与,才能将这种outside-in信号传递下去。研究发现,Toll样受体TLR的激活可诱导胞内MHCⅡ类分子与CD40以及Bruton酪氨酸激酶BTK结合,从而导致BTK持续活化以及TLR信号特异性基因转录的延长。除CD40外,还有研究报道B细胞受体复合体成分CD79a和CD79b、IgE受体以及CD19也参与了与MHCⅡ类相关的信号转导。通过MHCⅡ类分子进行信号传导仍然是一个相对较新的概念,这一现象在免疫调控中的真正后果,仍有待未来进一步揭示。
8 结论与展望
由于MHCⅠ类和MHCⅡ类抗原呈递通路在调控免疫反应中具有基础性作用,并且与移植、感染、疫苗以及自身免疫密切相关,围绕这两条通路的生物学研究已经开展得相当深入。尽管目前对抗原呈递过程的认识已经达到很高的细致程度,但从整体上看,这些认识其实仍只是整个系统的轮廓。要获得更深入的理解,还需要借助现代技术,例如siRNA筛选,从全基因组尺度系统考察相关分子之间的关系。这类方法能够为任何一个生物学过程提供较为完整的新分子清单。将siRNA数据与流式细胞术或显微成像数据结合,再整合定量PCR和微阵列获得的转录信息,已经使研究得以识别多条新的通路,并揭示新的GTP酶和马达蛋白在MHCⅡ类转运中的调控作用。这类实验数据集还可以继续与其他来源的数据整合,例如其他siRNA或遗传筛选结果,以及表达数据库和蛋白-蛋白相互作用数据库,从而在计算机中构建候选通路模型。不过,这些通路仍需要通过实验进一步验证,否则就可能给对MHCⅠ类和MHCⅡ类抗原呈递通路的理解引入噪声。
与此同时,对MHCⅠ类和MHCⅡ类抗原呈递的认识已经能够,而且事实上也正在被转化为治疗策略。随着对这些分子介导抗原呈递机制理解的不断加深,未来有望发现更多可用于治疗性调控免疫系统的新靶点。近来,许多研究团队已经揭示了抗原加工与呈递系统中的新步骤。然而,这一领域仍存在大量未知和争议,文中表2和表3结合MHCⅠ类与MHCⅡ类生物学领域专家的意见,对这些问题进行了总结。当前仍难以判断免疫优势肽是否可以被准确预测,也尚不清楚为什么某些特定的MHCⅠ类或MHCⅡ类等位基因会与自身免疫病相关。较为明确的例外是麸质、HLA-DQ2和HLA-DQ8与乳糜泻之间的已知联系。对于这些核心问题,未来几年有望逐步获得更清晰的答案。