Nat. Chem. Biol. 2026 | 两步式发条样机制开启 PIEZO2 的蛋白-脂质复合孔道
今天介绍的这项工作来自 nature chemical biology。该工作聚焦于机械敏感离子通道 PIEZO2 的门控机制这一长期未解的关键问题,结合混合分辨率分子动力学模拟与电生理实验,从结构与动力学层面系统揭示了力如何驱动通道开启。研究利用 PACE 力场,在接近生理张力条件下对全长 PIEZO2 进行长时间尺度(20 µs)模拟,克服了传统全原子模拟在时间与体系规模上的限制。结果表明,膜张力首先诱导外周跨膜“臂”结构发生扁平化,这一大尺度构象变化通过类似“钟表机构”的协同运动向中央孔道传递:伴随臂的重排,孔道相对于臂发生逆时针旋转,同时内侧孔道螺旋发生顺时针扭转,从而逐步打开离子通路。该过程呈现出明确的两步机制,包括一个低张力下稳定存在的次导通态(O1)以及完全开放态(O2),其中次导通态也得到了电生理实验的支持。进一步的全原子模拟显示,开放态孔道为蛋白-脂质共同构成的复合结构,其孔径、电导及离子选择性与实验结果高度一致。整体而言,该研究不仅在原子尺度上阐明了 PIEZO2 如何将膜张力转化为孔道开启,还为理解机械力在膜蛋白中的传递提供了普适性框架,同时也展示了混合分辨率模拟在研究大型膜蛋白动力学及药物靶点开发中的重要潜力。

获取详情:
0 摘要
机械敏感的 PIEZO 通道被认为是通过外周跨膜臂结构域在张力作用下发生“扁平化”而开启的,但其激活的结构基础仍不清楚。在该研究中,研究人员利用混合分辨率分子动力学模拟,揭示了在生理张力条件下,PIEZO2 臂的大尺度运动如何传导并转化为中央孔道中的细微门控运动。臂的扁平化与孔道相对于臂的逆时针旋转以及内侧孔道螺旋的顺时针扭转相关联。这些类似“钟表机构”的运动以两步方式打开孔道,形成完全导通状态以及一种在低张力下出现的稳定次导通状态,该状态已通过电生理实验检测到。完全开放的 PIEZO2 孔道由脂质和氨基酸共同构成,其结构再现了电生理测量中观察到的最小孔径、电导、离子选择性以及氯离子电流的外向整流特性。这些发现为理解 PIEZO2 的门控机制提供了结构层面的见解,并表明混合分辨率分子动力学是一种研究大尺度膜蛋白动力学以及指导药物发现的有力方法。
1 引言
哺乳动物的PIEZO通道包括PIEZO1和PIEZO2,在脊椎动物多种机械转导过程中发挥着关键作用,涉及细胞体积调控、神经元分化、上皮稳态维持、血管与淋巴系统发育、血压调节以及感觉生理等方面。PIEZO1几乎广泛存在于各种细胞类型中,而PIEZO2则主要表达于外周感觉神经元,负责将机械刺激转化为神经信号。由于其在多种生理与病理过程中的重要作用,PIEZO通道被认为是治疗炎症相关疼痛、胃肠道功能紊乱以及失禁等疾病的潜在药物靶点。
从结构上看,PIEZO通道由三个相同的大型亚基组成,每个亚基约包含2500个氨基酸残基。这些亚基共同形成一个域交换的中央孔道结构,其外围环绕着三条较长的跨膜结构域,通常称为“臂”或“叶片”。冷冻电镜解析的结构显示,这些臂在自然状态下呈现向上弯曲的构象,使整个蛋白形成类似倒置穹顶的形态。这种非平面结构会对周围脂质双层产生显著影响,使膜发生偏离平面的形变,从而形成较大的膜占据范围。
在该弯曲构象中,中央的离子通道被三条内侧孔道螺旋上的疏水性残基封闭,从而阻止离子的通过。多项实验结果表明,这些臂结构既能够感知膜的曲率变化,也能够响应膜的拉伸,从而在机械形变与孔道开启之间建立联系。相关研究通过在PIEZO1臂上引入构象探针并监测荧光信号,发现臂的运动与孔道开启在时间上具有明显相关性。进一步的冷冻电镜研究表明,当PIEZO1通道嵌入小型脂质体并处于outside-out构型时,由于膜与蛋白之间存在曲率不匹配,其臂结构会发生完全扁平化。利用最小荧光光子通量显微技术的研究则显示,渗透压诱导的膜拉伸与PIEZO1臂之间的空间分离存在相关关系,这一现象与张力诱导的臂扁平化机制相一致。此外,在一种持续活化的S2472E突变体结构中,其臂结构呈现部分扁平化状态,进一步支持了该机制的合理性。
尽管已有诸多研究进展,PIEZO通道中臂结构的大尺度扁平化运动如何传递至中央孔道并调控离子通透,仍缺乏清晰的机制解释。现有的PIEZO1多种构象结构虽然已被解析,但其孔道区域的空间分辨率较低,且缺失约三分之一的N端臂结构域。这一N端区域在PIEZO2结构中得到了完整解析,然而目前尚未获得PIEZO2在开放态或中间态下的结构或可靠模型,因此难以构建完整的通道门控机制。在这一背景下,全长PIEZO2结构为借助分子动力学模拟深入解析其门控机制提供了重要契机。
然而,全长PIEZO2膜蛋白体系的庞大规模使得在微秒时间尺度上研究其门控运动面临显著挑战,传统的全原子分子动力学方法难以胜任。为克服时间尺度限制,以往针对PIEZO1开放过程的全原子模拟通常需要施加非生理条件,例如施加约80 mN m
针对上述问题,研究采用了一种混合分辨率力场“Protein with Atomistic details in coarse-grained Environment”(PACE),该方法能够在粗粒化的Martini溶剂和膜环境中保留蛋白的原子级细节。与Martini蛋白模型不同,PACE模型无需依赖弹性网络约束来维持蛋白的二级和三级结构,因为氨基酸之间的重要各向异性相互作用,如氢键和π-π堆叠,能够以类似全原子模型的方式得到保留。已有研究表明,PACE力场的改进版本能够再现与全原子模拟结果高度一致的张力诱导PIEZO1开放态结构。在该研究中,通过对全长PIEZO2通道进行20 µs的PACE模拟,揭示了一个由张力驱动的两步开启路径,并通过电生理实验加以验证。这些协调一致、类似钟表运作的门控运动为理解机械敏感结构域如何将力传递至蛋白-脂质复合孔道并实现开启提供了新的结构层面认识。
2 结果
2.1 来自压力钳记录的PIEZO2拉伸敏感性
记录机械敏感通道拉伸激活电流的一种常用方法是通过在膜片钳电极背侧施加负压(吸力),即压力钳电生理技术。利用该方法,此前研究表明,与PIEZO1相比,PIEZO2表现出较低的拉伸敏感性以及减弱的失活特性。在该研究中,通过在转染的HEK293TPIEZO1KO细胞中记录PIEZO2的拉伸电流对上述结果进行了验证,这些细胞本身不表达内源性机械敏感通道(图1a,b)。由于在缺乏精确膜几何测量的情况下无法直接获得膜张力,因此通过将PIEZO2电流的实验半激活压力P1/2与两种已知T50的通道进行比较,来估算产生PIEZO2最大峰值电流一半所需的膜张力,即半激活张力T50。这两种通道分别为PIEZO1(T50=1.4 mN m
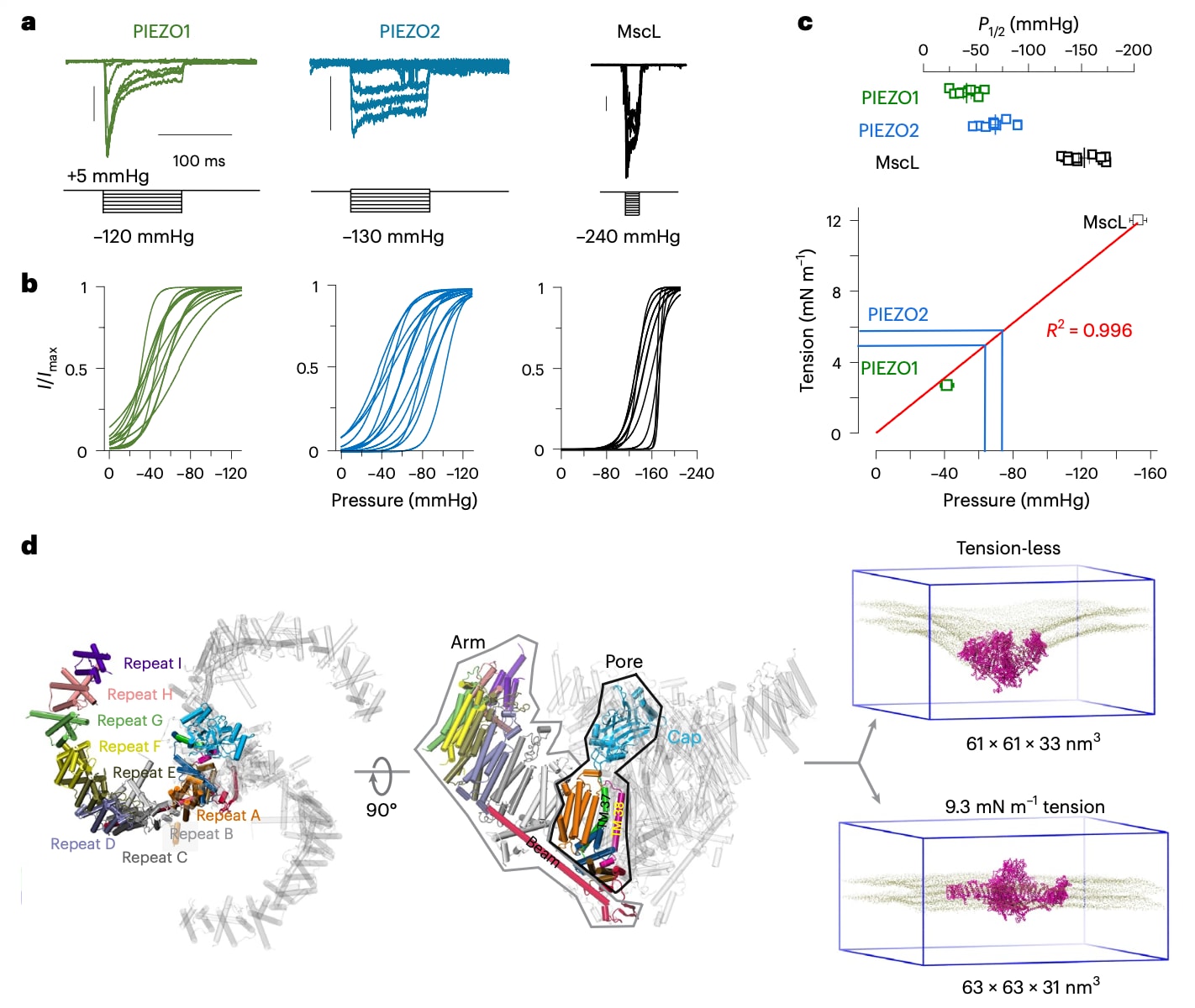
图1 | 模拟的 PIEZO2 系统与张力阈值。 a,代表性的拉伸激活电流轨迹:PIEZO1(左,绿色)和 PIEZO2(右,蓝色),记录电位为 −90 mV。每条曲线下方显示相应的移液管压力刺激方案。纵向标尺表示 100 pA。b,PIEZO1(绿色)和 PIEZO2(青色)的归一化拉伸激活(I/Imax),由实验数据通过玻尔兹曼理想化曲线拟合表示(数据可作为源文件获取;见“数据可用性”)。c,上:PIEZO1(绿色,n = 9)、PIEZO2(蓝色,n = 10)和 MscL(黑色,n = 11)的半激活压力 P1/2 的散点图。下:半激活张力与半激活压力之间的线性拟合(红线),PIEZO1(绿色方块)和 MscL(黑色方块)显示均值±标准误(s.e.m.)。蓝色线表示将 PIEZO2 的半激活压力从压力尺度换算到外推的张力尺度后的实验均值±标准误。d,左:PIEZO2 结构示意图,显示单个臂中的重复单元(A–I)。右:在 0 mN·m⁻¹ 和 9.3 mN·m⁻¹ 条件下模拟得到的平衡态 PIEZO2 构象(洋红色)及膜脂头基(棕褐色)。
2.2 膜尺寸与PACE模拟中的张力
利用优化后的PACE力场,对全长PIEZO2通道在约3700 nm
为了模拟膜拉伸,分别施加了0 mN m
2.3 模拟张力下PIEZO2孔径分布
在去垢剂环境中解析得到的PIEZO2结构(PDB:6KG7)显示其孔道处于关闭状态,并存在两个主要的收缩位点(扩展数据图2),其中一个由三个亚基中的L2743侧链构成,另一个由F2754的三个侧链形成。这些残基位于V2750的上下位置,V2750在PIEZO1中的对应残基为V2476,已被证实在全原子和PACE模拟中作为疏水性门控位点。通过在所有模拟中追踪这些残基侧链原子之间的平均最小距离,可以观察孔道的变化。在无张力条件下,孔道主要由L2743和V2750形成限制,而F2754的侧链在模拟过程中逐渐彼此分离。在施加张力的模拟中,孔道的主要收缩始终由V2750控制,其残基之间的侧链距离明显小于L2743和F2754(扩展数据图2)。值得注意的是,PIEZO2中的V2750与PIEZO1中的V2476均在通道失活过程中形成疏水性门控结构,提示这两个同源通道可能通过这一保守的缬氨酸残基动态调控离子通透。
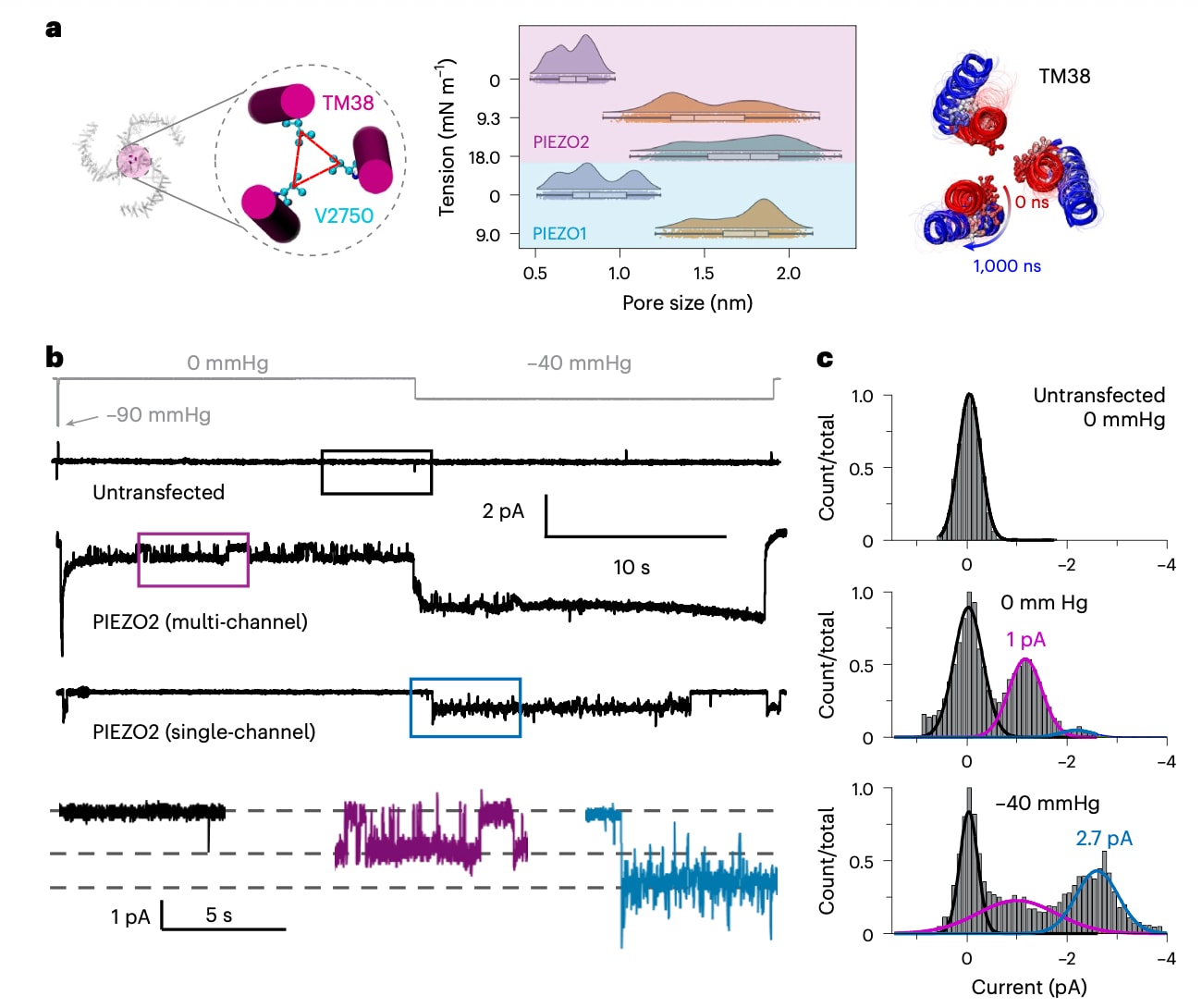
图2 | 张力模拟条件下 PIEZO1 和 PIEZO2 的孔径大小。a,左:通过测量 V2750 侧链之间的 Cβ−Cβ 距离来表征 PIEZO2 的孔径。中:在不同张力条件下模拟得到的 PIEZO1 和 PIEZO2 孔径分布。误差棒表示标准误(s.e.m.),基于以下重复计算:PIEZO2(0 mN·m⁻¹,4 次)、PIEZO1(0 mN·m⁻¹,3 次)、PIEZO2(9.3 mN·m⁻¹,9 次)、PIEZO2(18.0 mN·m⁻¹,7 次)以及 PIEZO1(9.0 mN·m⁻¹,4 次)。统计分析使用 500 ns 之后采样的构象。每个雨云图结合了概率密度分布,下方的箱线图显示中位数、四分位数和单个数据点。右:在 9.3 mN·m⁻¹ 条件下,0 ns(红色)与 1,000 ns(蓝色)时的 PIEZO2 TM38 重叠示例,其中 V2750 以 Corey–Pauling–Koltun 模型表示。b,上:未转染或转染 PIEZO2 的 HEK293TPIEZO1KO 细胞的压力钳记录示例。下:上述轨迹中各插图的放大视图。c,由四个独立膜片细胞汇总得到的全点直方图,并分别使用一个(上)或三个(中和下)高斯函数进行拟合。
因此,采用三个亚基中相邻V2750的Cβ-Cβ平均距离作为表征孔径大小的指标。在无张力模拟中,PIEZO2的平均孔径为0.72±0.05 nm,与冷冻电镜结构中关闭状态孔径0.66 nm一致(图2a及扩展数据表1)。在施加张力后,孔道发生扩张,并呈现两个稳定状态,对应孔径分别为1.36±0.02 nm和1.81±0.03 nm。尽管在两种张力条件下均可观察到这两种孔径状态,但其分布存在差异:在9.3 mN m
为了探究PIEZO1是否也表现出类似的孔径分布,进一步进行了约1 µs的PACE模拟,分别在0 mN m
2.4 PIEZO2亚导通状态的压力钳记录
为了在实验上验证PIEZO2是否存在次导通状态,对表达PIEZO2的HEK293TPIEZO1KO细胞进行了单通道电流记录。实验中将膜片电位保持在−90 mV,先施加一个短暂的−90 mmHg测试脉冲,随后在无吸力条件(0 mmHg)下记录20 s,再施加−40 mmHg的持续负压记录20 s(图2b)。在仅包含单个活性通道的膜片中,与−40 mmHg条件相比,0 mmHg下几乎不出现通道开启事件,这与PIEZO2在缺乏外部刺激时开启概率极低的特性一致。相比之下,包含多个通道的膜片在0 mmHg条件下也可观察到较为频繁的开启事件。
通过对单通道及多通道记录进行全点直方图分析,可以分辨出两种不同的单位电流幅值:在0 mmHg时约为1 pA,在−40 mmHg时约为2.7 pA,分别对应约11 pS和30 pS的两种电导(图2c)。其中较大的电导值与此前在加压膜片中测得的结果相近。这些实验结果与模拟观察相一致,表明PIEZO2在低张力条件下更倾向于处于次导通状态,而在较高张力条件下则主要处于完全导通状态。
2.5 张力下孔道中的水和脂质占据情况
PIEZO2的孔道在胞内侧对称地分叉形成三个侧向开口,这一结构特征与此前对PIEZO1的观察一致(扩展数据图3a)。在各条模拟轨迹中,对孔腔内水分子、脂质尾部以及脂质头基原子的动态分布情况进行了分析(扩展数据图3b及补充图4)。在无张力条件下,仅有少量水分子分布在由三个V2750侧链构成的疏水收缩位点的上下区域。随着张力诱导孔道扩张,孔腔内水分子和脂质头基的数量随之增加,同时脂质尾部被排除在孔道之外,这一变化在时间上与孔径扩大的过程相一致。
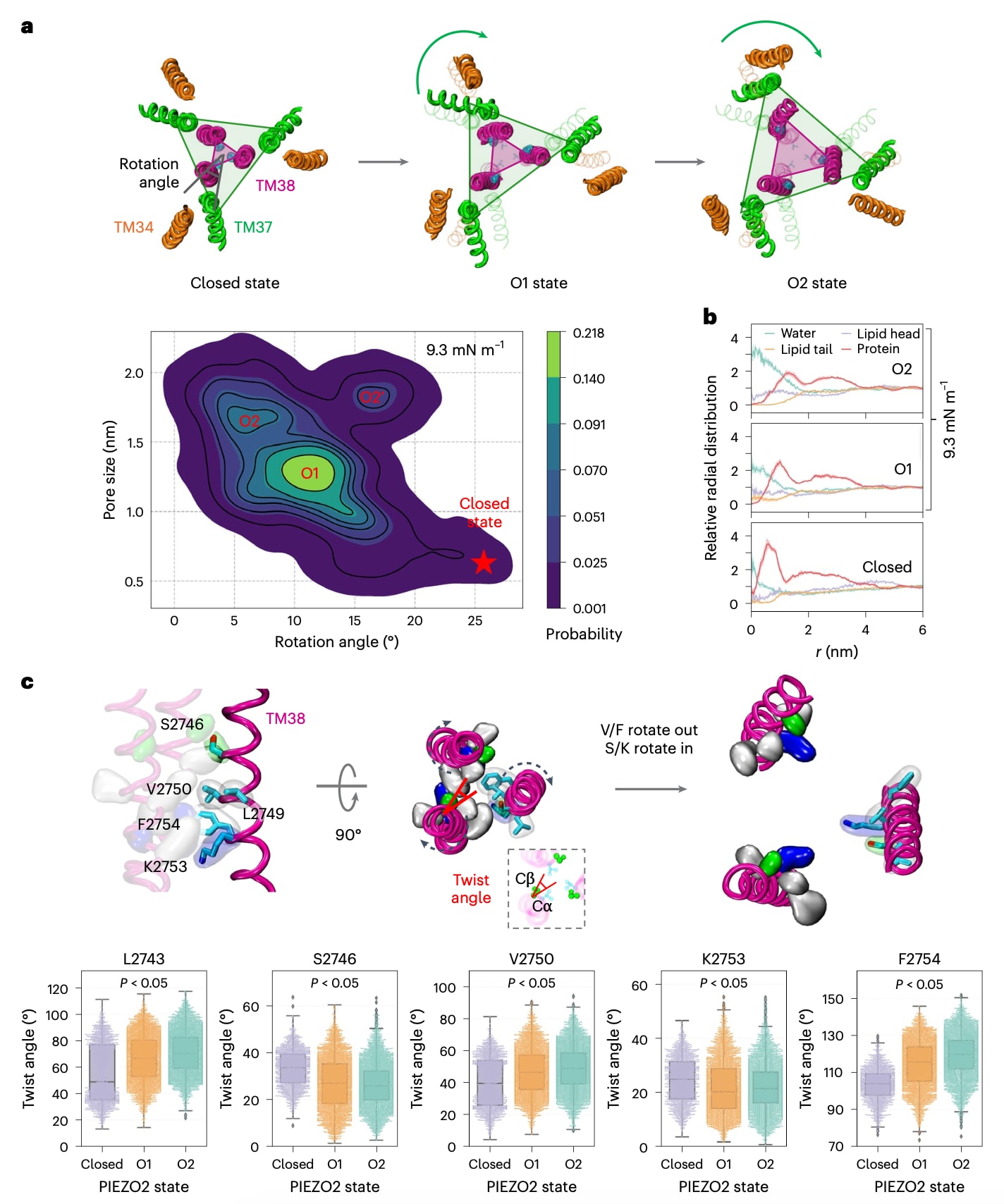
图3 | 时钟式 PIEZO2 门控运动。 a,上:孔道螺旋的门控运动。下:内外三角之间的相对夹角(分别由 TM38 和 TM37 三聚体构成)与孔径大小的关系,数据来自 9.3 mN·m⁻¹ 条件下的全部 9 个重复(补充视频 2)。密度分布通过核密度估计计算(补充图 5)。b,脂质头部、脂质尾部、水和蛋白的径向分布函数(RDF)(孔中心定义为三个 V2750 的 Cα 原子的质心)。闭合状态分析了 0 mN·m⁻¹ 条件下的全部 4 个重复。9.3 mN·m⁻¹ 条件下的 9 个重复被分类为 O1 和 O2 两种状态,并分别计算各自的 RDF(阴影区域表示标准误,s.e.m.;扩展数据图 4)。c,上:TM38 的顺时针扭转及侧链重定位。插图:扭转角定义为从 Cα 指向 Cβ 的 xy 向量与从 Cα 指向孔轴(所有 TM38 的质心)的 xy 向量之间的夹角。V,缬氨酸;F,苯丙氨酸;S,丝氨酸;K,赖氨酸。下:极性和疏水性 TM38 残基在闭合状态(0 mN·m⁻¹,淡紫色,n = 4)、O1(橙色,n = 9)和 O2(薄荷色,n = 10)中的扭转角。箱线图显示中位数和四分位数,蜂群图展示单个数据点。误差棒表示由每个重复的平均扭转角计算得到的标准误(s.e.m.)。不同状态之间的扭转角差异通过单因素方差分析(one-way ANOVA)进行检验,使用所有数据点(闭合态 1,842;O1 3,703;O2 4,824),其中 L2743(P = 3.39 × 10⁻¹⁸)、S2746(P = 4.57 × 10⁻¹⁹)、V2750(P = 2.46 × 10⁻¹²)、K2753(P = 2.56 × 10⁻⁵)和 F2754(P = 5.98 × 10⁻¹⁸)显示出统计学显著性。仅分析了每条轨迹中 500 ns 之后采样的构象。
2.6 PIEZO2开启的钟表式门控运动
在所有施加张力的模拟中,原本具有弯曲构象的PIEZO2臂均发生扁平化,这一现象与以往多项研究的预测一致。然而,这种扁平化如何触发孔道开启一直缺乏清晰解释。模拟结果显示,臂的扁平化会诱导内侧孔道螺旋(TM38)相对于外部结构发生逆时针旋转。通过计算由TM38构成的内三角形与由TM37构成的外三角形之间的相对角度,可以量化这一旋转运动(图3a)。在9.3 mN m
除内外孔道螺旋之间的相对旋转外,孔道开启还伴随着每条内侧螺旋围绕自身α螺旋轴的顺时针扭转(图3c)。通过计算各残基Cα-Cβ连线与Cα-孔轴连线在xy平面的夹角(即扭转角),并对三个亚基取平均,可以判断侧链是朝向还是远离孔腔移动。结果显示,包括L2743、V2750和F2754在内的大多数非极性侧链的扭转角增加,表明其逐渐远离孔腔;而极性残基S2746和K2753的扭转角减小,说明其重新朝向孔腔取向。这种协同运动使孔道收缩区域的化学环境在物理扩张过程中由疏水性转变为亲水性,从而进一步促进孔道的去脂化和水化过程。同时,在O2状态下TM38的扭转程度较O1更为显著,提示该扭转运动与形成更大且更亲水的孔道相关。
为了进一步验证这些开放态的稳定性,在PACE模拟得到的O1、O2和O2′构象基础上,对孔道区域(包括Repeat A、帽结构、孔道及C端结构域)进行了两组独立的100 ns全原子模拟(扩展数据图5)。结果表明,O1和O2状态下的孔径在时间上保持较为稳定,尽管内外螺旋之间的旋转角略有减小(约7°);而较少出现的O2′状态则会逐渐转变为O1或O2状态。采用CHARMM36m力场的全原子模拟结果支持PACE模拟中观察到的这两种稳定开放态。
2.7 PIEZO2穹顶形态在张力下的变化
进一步对PIEZO2从关闭态到O1和O2状态过程中穹顶形态的变化进行了量化分析。在去垢剂环境中解析得到的全长PIEZO2结构中,三条臂的跨膜区域呈现出向上弯曲的倒置穹顶形态,其臂角(臂与PIEZO内部轴之间的平均夹角)为53°(图4a)。以三条内侧螺旋TM38和最外侧重复结构的几何中心作为参考点,穹顶高度为8.5 nm,胞外帽结构的高度为6.0 nm(图4b),而由三条臂末端界定的穹顶在膜平面上的投影面积Aproj为385 nm
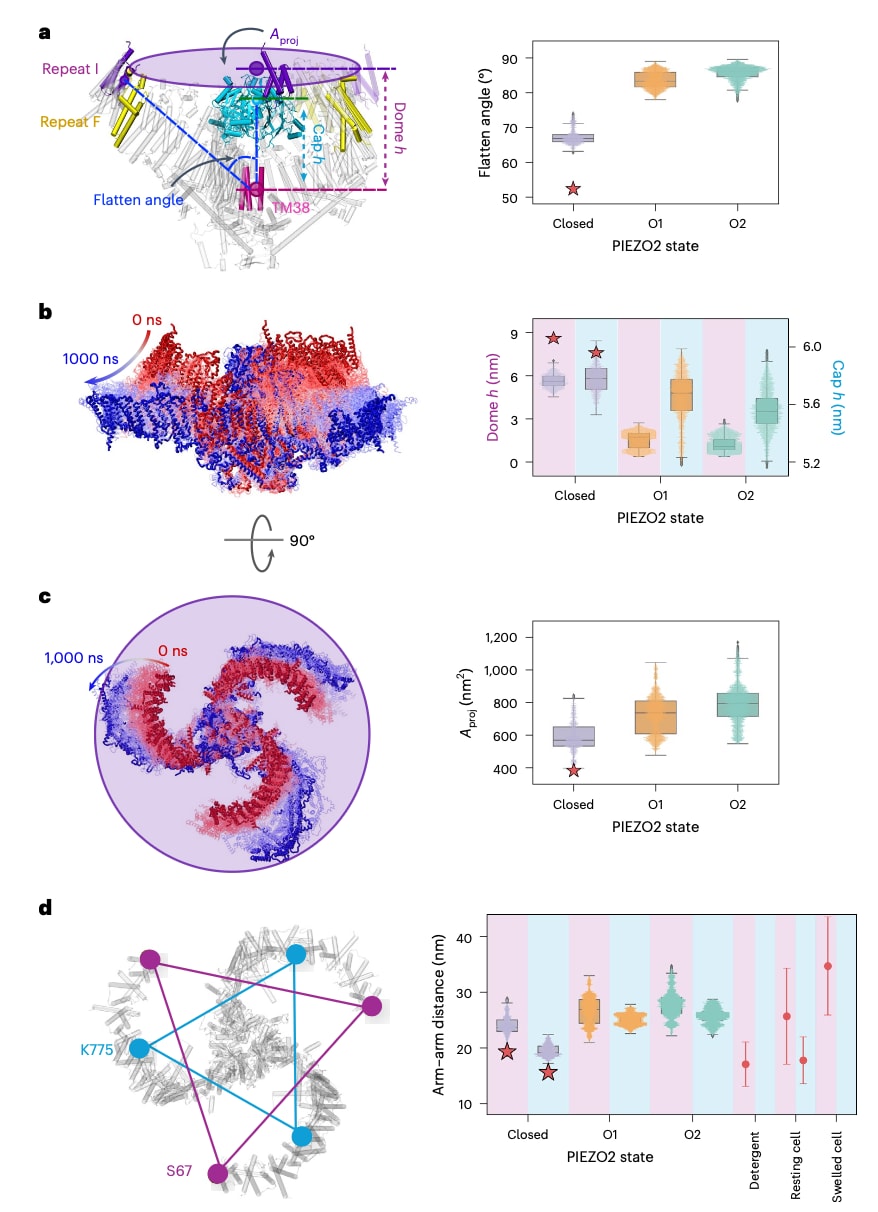
图4 | 张力诱导的 PIEZO2 全局构象变化。 a,臂角定义为臂轴(由最后一个重复单元 Repeat I 的质心确定)、内侧孔道螺旋(TM38)以及 PIEZO2 内部轴(由内侧三条孔道螺旋和帽区的质心确定)之间夹角的平均值。b,穹顶高度(h)定义为三个 Repeat I 与三个 TM38 在 z 轴方向上的质心距离。帽区高度(cap h)定义为帽区与三个 TM38 在 z 轴方向上的质心距离。洋红色和青色分别表示穹顶高度和帽区高度。c,穹顶投影面积为 Aproj = π × r² = π × (a×b×c) / √[(a+b+c)×(a+b−c)×(a−b+c)×(b+c−a)],其中 r 为投影半径,a、b、c 为三个 Repeat I 质心之间的距离。d,残基 S67(洋红色背景)和 K775(青色背景)之间的臂间距离变化图,对应于 PIEZO1 的 Q103 和 D670。右:在不同条件下(去垢剂中、静息细胞或膨胀细胞),通过 MINFLUX 测量得到的 PIEZO1 Q103 和 D670 的臂间及外臂距离(参考文献 13)。箱线图中,水平线表示中位数,四分位距(IQR)范围为第一至第三四分位数(Q1 至 Q3),须线延伸至 1.5×IQR 范围内(超出该范围的点为离群值)。所有数值均以均值±标准误(s.e.m.)表示。分析中排除了每条分子动力学轨迹前 500 ns 采样的构象。对 9.3 mN·m⁻¹ 和 18.0 mN·m⁻¹ 条件下共 16 个重复的 PIEZO2 孔径分布进行了核密度估计(KDE)分析,并以主局部最小值 1.54 nm 作为阈值,将状态定义为 O1(≤1.54 nm)和 O2(>1.54 nm)。该分析包括 0 mN·m⁻¹ 条件下的 4 个重复,以及 O1 的 9 个重复和 O2 的 10 个重复。在每种情况下,红色星号表示在去垢剂胶束中解析得到的 PIEZO2 冷冻电镜结构(PDB:6KG7)的对应数值。
当该PIEZO2模型嵌入无张力的Martini POPC双层膜中时,该膜的弯曲刚度为25.2±2.2 kBT,其中kB为玻尔兹曼常数,T为温度。在这一条件下,穹顶高度和帽结构高度分别降低并稳定在5.5±0.1 nm和5.8±0.1 nm,同时臂角增大至67.8±1.0°,Aproj扩展至590±51 nm
将PIEZO2穹顶近似为球冠结构时,以最外侧Repeat I的几何中心为参考,其曲率半径在关闭态、O1态和O2态下分别为20±2 nm、101±18 nm和122±14 nm。然而,由于外侧重复结构I、H、G和F具有一定柔性,PIEZO整体并非理想的球冠,在边缘区域会出现一定程度的扁平化。以更靠近中心的Repeat E作为参考时,测得的曲率半径系统性较小,在关闭态、O1态和O2态下分别为16±1 nm、70±12 nm和88±11 nm。为了与多个仅包含部分臂结构的PIEZO1冷冻电镜结构及分子动力学模拟结果进行比较,同样采用Repeat E对穹顶形态进行测量。总体来看,模拟结果与多种实验结构数据表现出良好一致性。
2.8 PIEZO2在张力下的臂间距离
MINFLUX技术已被用于在细胞水平测量PIEZO1的整体构象变化。由于PIEZO1与PIEZO2在结构上的高度相似性,这些实验数据可以与分子动力学模拟中得到的整体结构变化进行直接比较。基于这一思路,对不同张力条件下PIEZO2的臂间距离进行了测量(图4d及扩展数据表1)。
在MINFLUX实验中,PIEZO1中Repeat I区域Q103残基之间的平均臂间距离在去垢剂环境中为17.1±4.0 nm,在静息细胞膜中为25.7±8.6 nm,而在渗透胀大的细胞中达到34.7±8.8 nm。模拟结果显示,PIEZO2中对应的Repeat I区域S67残基之间的距离呈现相似变化趋势,从无张力双层膜中的23.8±1.1 nm增加到O1状态的26.8±0.7 nm和O2状态的27.6±0.6 nm。对于Repeat F区域,MINFLUX测得PIEZO1中D670残基之间的距离在静息细胞膜中为17.8±4.2 nm,而模拟中PIEZO2对应的K775残基之间距离在无张力条件下为19.6±0.5 nm,在O1和O2状态下分别增加至25.2±0.3 nm和25.7±0.4 nm。
近期研究表明,PIEZO1臂的延展程度还会受到亚细胞结构、细胞骨架完整性以及化学调节因子的影响。总体来看,分子动力学模拟中在PIEZO2两个臂位置测得的三角化距离均落在多个MINFLUX实验结果的标准差范围内,从而为模拟中捕捉到的整体构象变化提供了重要的实验支持。
2.9 臂运动与孔道扩张之间的耦合
为阐明臂的大尺度运动如何引发内侧孔道螺旋的小尺度重排,首先对9.3 mN m
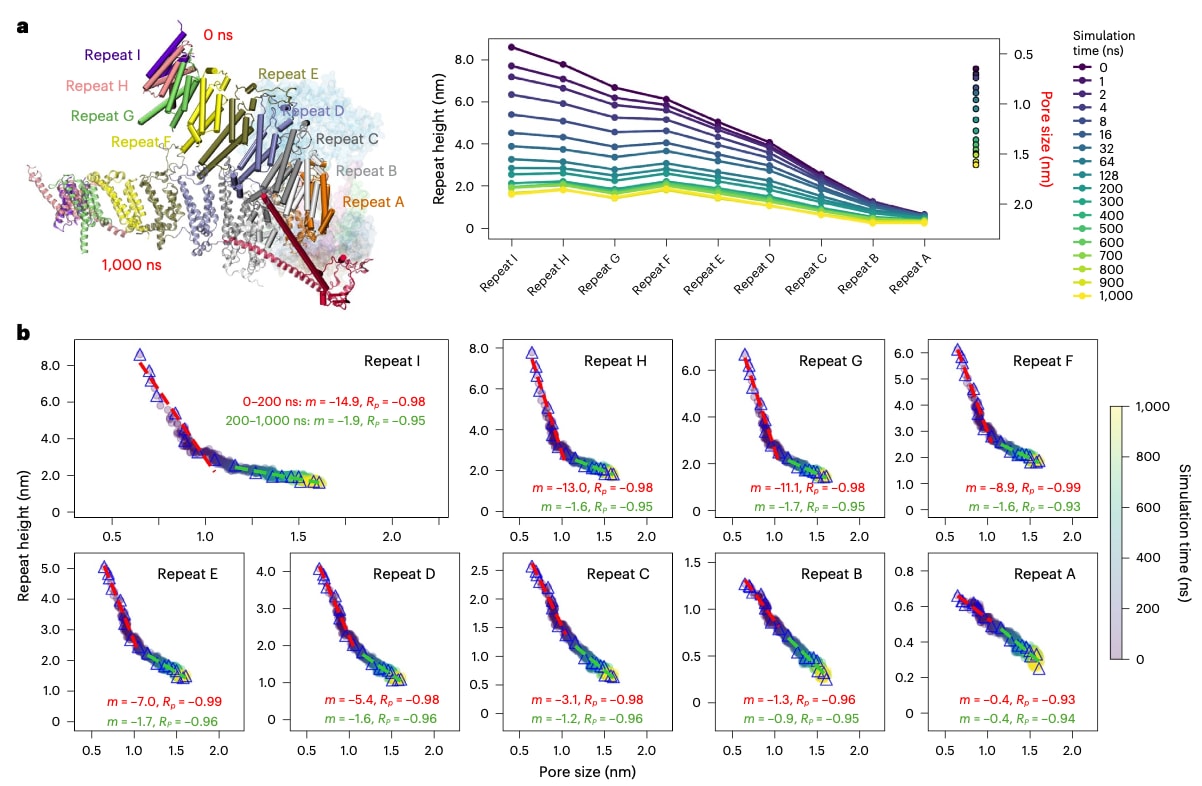
图5 | 穹顶形状与孔径之间的耦合关系。 a,左:单个臂在 0 ns 和 1,000 ns 时的结构快照。右:每个跨膜螺旋重复单元的穹顶高度随时间的变化。各面板表示在 9.3 mN·m⁻¹ 张力条件下的模拟结果。折线图显示从最外侧的 Repeat I 到中心的 Repeat A 的所有重复单元在模拟过程中的平均高度变化。高度通过在相同时间点、相同张力条件下对多个重复(replicas)的结果取平均得到。孔径以散点图形式表示。b,在 9.3 mN·m⁻¹ 条件下,重复单元高度与孔径随时间变化的关系。为捕捉穹顶高度初始快速下降及随后趋于稳定的过程,选取了一系列特定时间点(0、1、2、4、8、16、32、64、128、200、300、400、500、600、700、800、900 和 1,000 ns;蓝色三角),在下降阶段采用更密集的采样,而在系统接近稳态时采用较稀疏的采样。基于这些数据,对前 200 ns(红线)和随后 200–1,000 ns(绿色线)分别进行了线性拟合。0 mN·m⁻¹ 和 18.0 mN·m⁻¹ 条件下的分析见扩展数据图 7 和 8。
2.10 PIEZO2电导与整流特性的全原子模拟
由于PACE模型采用的Martini水和离子模型不适合用于计算动力学性质,因此利用全原子水平的PIEZO2 O1和O2孔道模型对单位电导进行了估算。与PACE模拟结果一致,全原子模拟显示相邻内侧孔道螺旋之间的间隙被POPC脂质填充(图6a)。在施加恒定电压的条件下,观察到类似于此前PIEZO1模拟中的多窗口离子传导通路。胞外帽结构的下部对水分子和离子是可达的,而其顶部仍保持封闭状态(图6b)。通过统计离子穿过孔道的数量可以发现,O2状态的电导显著高于O1状态,在−0.6 V和+0.6 V条件下分别表现为34.7±2.7 pS和38.2±7.7 pS,而O1状态分别为8.9±4.1 pS和11.6±6.2 pS(扩展数据表2),这一结果与实验测量值(约11 pS对应0 mmHg,约30 pS对应−40 mmHg)高度一致(图2b,c)。此外,O2状态下的计算电导也与已报道的小鼠PIEZO2电导值相符。值得注意的是,在O2状态下,外向电流(+0.6 V)中氯离子的贡献大于内向电流(−0.6 V),这一结果与近期实验观察一致,即哺乳动物PIEZO通道在加压膜片中表现出氯离子电流的外向整流特性,而这一特性在O1状态中并不明显。
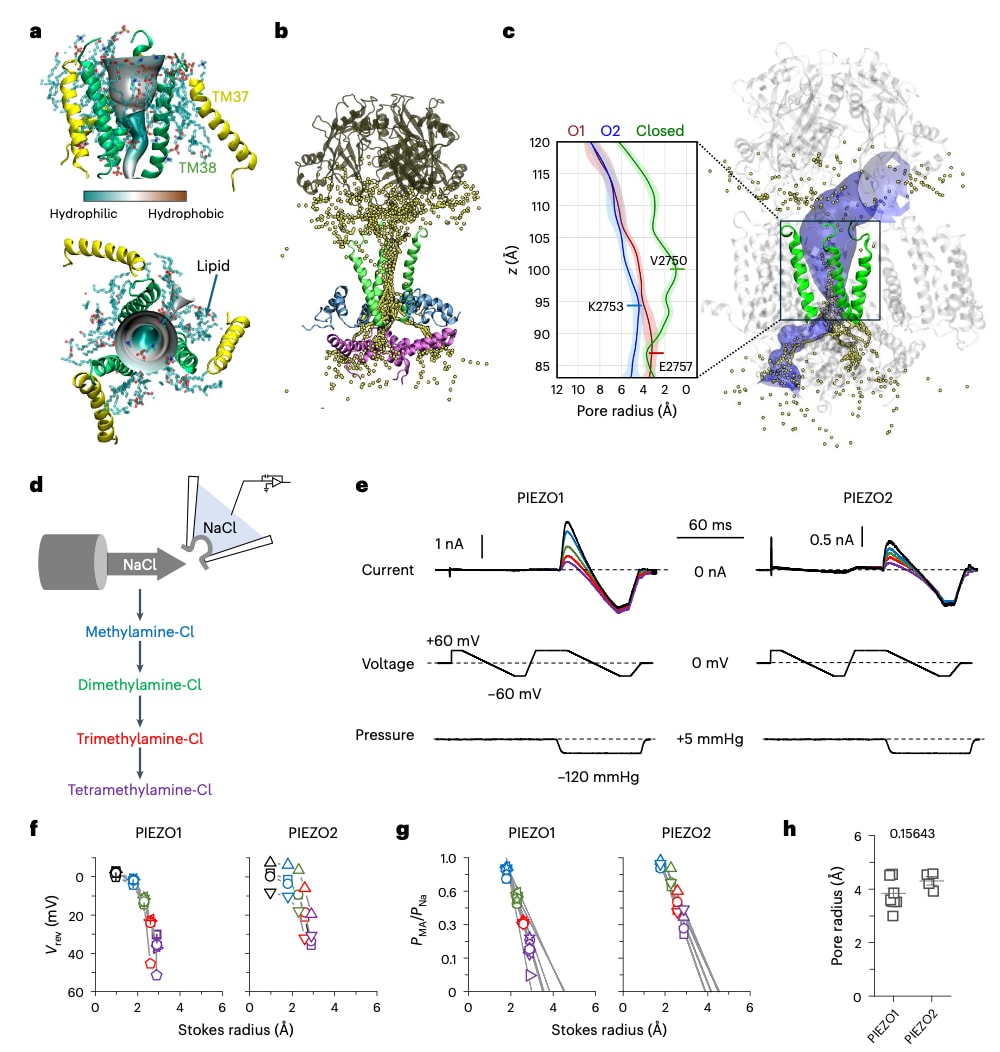
图6 | PIEZO2 开放孔的全原子模拟及最小孔半径。 a,PIEZO2 完全开放孔(O2 状态),显示外侧孔道螺旋(TM37,黄色)和内侧孔道螺旋(TM38,绿色),并与以棒状模型表示的 POPC 脂质相互嵌合(青色为碳,红色为氧,蓝色为氮,棕褐色为磷)。三维孔道形状按亲水性着色,使用 CHAP 生成。b,在 +0.6 V 条件下进行 100 ns 模拟所得的 K⁺ 离子重叠坐标。蛋白主链显示为帽区(棕褐色)、TM38(绿色)、部分锚定结构域(残基 2,385–2,435,蓝色)以及 C 端结构域(残基 2,770–2,822,洋红色)。c,使用 HOLE 计算的闭合态(绿色)、O1(红色)和 O2(蓝色)状态的孔径半径分布曲线。均值及标准差(阴影部分)基于从总计 300 ns 中提取的 40 个帧计算得到。右:O2 状态的代表性三维孔道轮廓(蓝色表面)及 K⁺ 渗透路径(黄色点)。d,对切除膜片进行细胞内盐溶液的顺序灌流。e,根据 d 中所示方案获得的 PIEZO1 和 PIEZO2 电流轨迹示例。f,g,散点图表示不同甲胺相对于 Na⁺ 的反转电位(f)和通透性(g),并作为各测试甲胺的 Stokes 半径的函数,分别对应 PIEZO1(独立膜片数 n = 9)和 PIEZO2(独立膜片数 n = 4)。h,散点图显示通过线性外推计算得到的 PIEZO1 和 PIEZO2 开放孔的最小半径(图上方数字为双尾非配对 Student t 检验的精确 P 值)。f–h 中的误差棒表示标准误(s.e.m.)。
2.11 PIEZO2孔径的电生理测量
利用HOLE程序对关闭态、O1和O2状态的孔径分布进行分析发现,最小孔径沿通道逐渐增大,从V2750位置的1.06 ± 0.29 Å增加至K2753位置的4.38 ± 0.57 Å(图6c)。为了验证这些开放态孔道尺寸,通过电生理方法测定较大分子通透物的相对通透性,从而估算PIEZO1和PIEZO2的最小孔径。
具体方法是测量切除膜片中拉伸诱导的宏观电流反转电位,实验从对称的NaCl溶液开始,随后转为非对称条件,即电极中为NaCl,浴液中依次替换为尺寸逐渐增大的有机阳离子(甲胺、二甲胺、三甲胺和四甲胺)(图6d,e)。在对称与非对称条件下获得的反转电位用于计算甲基化胺相对于钠离子的通透性(PMA),并对氯离子的通透贡献进行校正。随后将这些通透性数据与对应分子的已知Stokes半径进行关联分析。
基于排除体积理论对上述关系进行线性回归拟合,从而计算最小孔径(图6f-h)。结果显示,PIEZO1和PIEZO2的最小孔半径分别为4.21 ± 0.17 Å和4.35 ± 0.16 Å,二者在统计学上无显著差异(P = 0.58925),且与此前报道的PIEZO1孔径以及该研究中完全导通的PIEZO2开放孔径结果一致。
3 讨论
该研究构建了一个描述PIEZO2在膜拉伸作用下激活过程的计算模型,并通过电生理实验对张力敏感性、电导、离子选择性、整流特性以及最小孔径等多个方面进行了系统验证。为克服大型蛋白-膜体系建模中的固有挑战,采用了优化后的PACE混合力场。与此前针对PIEZO1的全原子模拟不同,PACE方法能够将通道嵌入到约3700 nm
支持两步“钟表式”门控机制的重要证据在于,在低张力条件下PIEZO2主要处于次导通状态,这一现象在电生理记录和模拟中均得到一致观察。在较高张力下,随着TM38螺旋进一步分离,孔道扩展为完全导通状态。这种行为在离子通道中相对少见,因为尽管次导通状态较为常见,但通常稳定性低于完全导通状态。在PIEZO1的PACE模拟中,即使在低张力下次导通状态的占比也较低,这一结果与单通道记录中观察到的短暂次导通状态相一致。
关于保守残基V2750在激活过程中作为疏水性门控位点的观察,并不与其在PIEZO2失活过程中同样作为门控位点的观点相矛盾。从广义上看,激活门是指离子通透路径中的物理收缩位点,其消失是离子传导发生的前提;而失活门则是在持续刺激下形成的收缩结构,用于终止离子传导。在某些离子通道中,这两类门可能位于不同位置,例如钾通道中激活门位于胞内螺旋束交汇处,而C型失活门位于选择性滤过器处,但并无证据表明二者必须在空间上分离。该研究获得的PIEZO2开放态结构模型可为未来解析其失活机制的结构基础提供重要参考。
由于PIEZO呈三聚体结构,孔道扩张过程中内侧螺旋的分离不可避免地会在螺旋之间形成较大的间隙,从而为POPC脂质进入孔腔提供通道。然而,在该研究中观察到的TM38螺旋扭转运动会将孔道的化学环境由疏水性转变为亲水性,从而促进孔腔水化。这种化学环境的变化能够阻止POPC脂质进入孔腔并堵塞通道。同时,这也提示体相脂质可能作为开放态孔壁的组成部分存在,因为POPC脂质在结构上始终被相邻的内侧孔道螺旋夹持。类似的蛋白-脂质复合开放孔结构也在OSCA/TMEM63通道家族中被观察到。
已有研究提出,体相脂质通过膜侧窗在通道孔道内的进出,可能参与某些机械敏感离子通道的张力依赖性门控过程,例如OSCA/TMEM63、MscS以及TRAAK通道。特别是同源三聚体的酸敏感离子通道ASIC3在开启时也会形成类似的间隙,允许POPC脂质部分进入孔道并降低单通道电导。尽管先前的计算研究曾提出PIEZO1孔道可能被脂质完全阻塞,但近期研究表明,模拟中孔道内脂质密度可能因水化程度被低估而出现高估现象。该研究提出的“钟表式”门控机制并不支持PIEZO2主要通过脂质进出孔腔来感知膜张力的观点,但并不能排除其他类型脂质通过占据孔道而调节通道功能的可能性。事实上,多种脂质已被证实能够显著调节PIEZO1和PIEZO2通道功能,包括胆固醇、膳食脂肪酸、磷脂酸、磷脂酰肌醇类、神经酰胺以及磷脂酰丝氨酸等。此外,近期解析的人源PIEZO1与脂质修饰调控蛋白复合物结构中也在孔道内观察到脂质密度。基于PIEZO1和PIEZO2开放态结构开展包含调控脂质的全原子模拟,将有助于进一步区分脂质调控是否来源于膜力学性质的改变或特异性脂质-蛋白相互作用。
通过小分子调控PIEZO2通道活性被认为是治疗多种疾病的潜在策略,包括炎症相关疼痛、胃肠道疾病以及失禁等。目前尚无已知对PIEZO2具有选择性的抑制剂或激动剂,但开放态结构模型及其门控机制为基于结构的药物设计提供了重要基础。类似策略已在PIEZO1研究中成功应用,例如在Repeat A与Repeat B之间鉴定出别构激动剂结合位点并发现新的通道激活剂。该研究进一步发现,PIEZO2臂与孔道之间的机械耦合在最内侧的Repeat A处最强,因此推测位于Repeat A与B之间的小分子也可能对PIEZO2功能产生调控作用。
该研究仍存在一定局限性。首先,PIEZO2可能通过不同机制感知不同类型的机械刺激。例如,其胞内无序结构域对于由细胞骨架传递的力的激活是必需的,但对于膜拉伸诱导的激活则并非必需。该研究未包含胞内结构域和细胞骨架成分,因此主要揭示的是膜拉伸条件下孔道收缩区域的开启机制。此外,PIEZO1的结构研究表明,除膜孔门之外,通道中还可能存在其他通透门,例如位于胞内侧窗的侧向门。由于这些附加门的结构基础在PIEZO2中尚未解析或缺失,因此未能对其在拉伸激活中的作用进行研究。最后,尽管PIEZO1与PIEZO2在张力诱导下表现出相似的门控运动,如孔道扩张和穹顶扁平化,但PIEZO2需要更高张力才能开启的原因仍不清楚。结构比较显示,在关闭态下PIEZO1孔道上部区域较PIEZO2更加扩张,这可能有助于其更易开启。关于两者在膜张力响应上的差异机制仍需进一步实验研究加以阐明。
4 方法
4.1 PIEZO2原子模型的构建
PIEZO2模型基于小鼠全长PIEZO2在关闭状态下的冷冻电镜结构(PDB:6KG7)构建。PIEZO2同源三聚体包含114条跨膜螺旋,每个单体由38条跨膜螺旋组成。在PIEZO2结构中未解析的部分中,有三个胞外环区域(分别为Repeat C或THU7中的残基1383−1426,Repeat B或THU8中的残基2056−2074以及Repeat A或THU9中的残基2283−2298)与PIEZO1中对应区域具有超过60%的序列相似性。由于PIEZO1的这些区域在多种构象和不同分辨率下已有结构解析,因此选取PIEZO1结构(PDB:6B3R和7WLU)作为模板,利用MODELLER 10.4程序进行同源建模。此外,模型中还缺失连接帽结构与外侧和内侧孔道螺旋(TM37和TM38)的连接环区域(残基2493−2500和2732−2738),这些部分同样通过MODELLER中的环建模模块进行构建。同时,还对连接beam结构C端与latch区域的43个残基(1515−1557)进行了建模。最终得到的PIEZO2模型每个单体包含1953个残基(8−79,122−142,201−304,332−358,491−615,675−823,934−1577,1668−1727,1948−2111和2236−2822),整个三聚体共包含5859个残基。其余主要为缺乏模板结构的胞质无序区域未被建模。随后使用PDBFixer程序对模型中缺失的重原子进行修复。
4.2 穹顶形膜结构的构建方法
在此前针对截短PIEZO1冷冻电镜结构的分子动力学模拟中,通常将PIEZO1蛋白嵌入平坦的全原子或Martini脂质双层中,并在长时间平衡模拟及蛋白骨架约束条件下,膜会自发弯曲形成包裹蛋白的穹顶结构。然而,对于全长PIEZO2关闭态结构,其外围臂的跨膜区域位于胞外帽结构之上,形成更深的穹顶形态,难以通过简单的长时间模拟实现膜的自发包裹。为解决这一问题,开发了一种方法以高效构建能够完全包裹PIEZO2穹顶结构的膜拓扑。
首先,根据PIEZO2穹顶的几何形状,构建了一个由2129个虚拟粒子组成的半球结构以及一个小半径为2 nm、大半径为16 nm、由1800个粒子组成的环面结构。这两个结构沿z轴方向相距16 nm放置。随后,利用insane.py脚本构建了一个尺寸为60 nm × 60 nm × 44 nm的平坦Martini2 POPC脂质双层膜,并将半球和环面嵌入该体系中,其中半球位于脂质层上方,环面位于下方。通过引入Lennard-Jones势(C6 = 0,C12 = 0.01),在虚拟粒子与Martini POPC粒子之间施加排斥相互作用。
最后,固定环面的位置,以z轴为反应坐标,施加100000 kJ mol
4.3 PIEZO2粗粒化Martini体系的构建与模拟
首先将原子级PIEZO2结构转换为标准的粗粒化Martini2弹性网络模型(ELNEDYN22),采用0.9 nm的成对距离截断以及500 kJ mol
4.4 PIEZO2 PACE体系的构建与模拟
基于原子级PIEZO2模型构建其统一原子结构及参数,采用针对膜蛋白模拟优化并在PIEZO1模型中验证过的PACE力场。随后通过对齐蛋白骨架粒子,将已平衡的Martini体系中的蛋白替换为PACE模型,同时保留原有的Martini膜和溶剂环境。PACE体系包含5859个蛋白残基、11355个POPC脂质、896114个粗粒化水粒子、11390个Na
在PACE力场优化中发现,将跨膜螺旋主链氢键强度提高至1.5倍或2倍可维持膜蛋白结构稳定(整体r.m.s.d.分别为3.24 ± 0.12 Å和3.30 ± 0.05 Å),并与全原子模拟在螺旋区域氢键数量上具有良好一致性。鉴于螺旋结构完整性对于机械转导至关重要,因此在该研究中采用膜区螺旋氢键强度增强2倍的PACE参数进行PIEZO2模拟。
所有PACE模拟同样使用GROMACS(2021和2022版本)。Lennard-Jones相互作用和库仑相互作用的截断距离均设为1.2 nm,长程静电相互作用采用反应场方法处理。体系温度通过Nose−Hoover恒温器维持在323 K,压力通过Parrinello−Rahman压强耦合在半各向同性条件下控制为1 bar,压缩率为3 × 10
体系首先进行5000步能量最小化,随后进入1.2 ns预平衡阶段,在此过程中对蛋白Cα原子施加强位置约束(500 kJ mol
为了研究膜张力与通道激活之间的关系,对PIEZO2 PACE体系中的膜平面(xy方向)施加三种不同压力条件:+1 bar(对应无张力膜),−2 bar(对应9.3 ± 0.2 mN m
4.5 PIEZO1 PACE体系的构建与模拟
PIEZO1的PACE模型基于此前构建的全原子模型,该模型来源于冷冻电镜结构(PDB:6B3R)。每条PIEZO1链被划分为四个独立片段:片段I(残基782–1365,包含Repeat C–F及beam结构),片段II(残基1493–1578,clasp结构),片段III(残基1655–1807,Repeat B),以及片段IV(残基1952–2546,包括Repeat A、anchor、TM37、cap、TM38及C端结构域)。
在体系构建过程中,首先利用CHARMM-GUI Martini Maker将PIEZO1的粗粒化模型嵌入到尺寸为50 nm × 50 nm × 35 nm的Martini POPC脂质双层中。实验结构被转换为ELNEDYN粗粒化模型,并在对骨架粒子施加谐约束(力常数为1000 kJ nm
为了研究膜张力对PIEZO1孔道开启的影响,在PACE模拟中对膜平面施加两种不同的侧向压力:+1 bar(对应0 mN m
4.6 PIEZO2全原子体系的构建与模拟
基于PACE模拟得到的O1和O2开放态模型,采用CHARMM36m力场构建PIEZO2的全原子模型。仅保留中央核心结构域(包括Repeat A、cap、孔道及C端结构域),并将其嵌入POPC脂质双层中,同时在0.15 M KCl溶液中进行溶剂化。O1开放态体系包含465,763个原子,其中包括121,269个水分子和543个POPC脂质分子;O2开放态体系包含474,090个原子,其中包括124,084个水分子和542个POPC脂质分子。
全原子模拟使用GROMACS(2023.3版本)进行。范德华相互作用采用1.2 nm的截断距离,并在1.0 nm至1.2 nm范围内使用力切换函数。长程静电相互作用通过粒子网格Ewald方法计算,短程静电截断距离为1.2 nm。所有含氢键的化学键通过LINCS算法约束,时间步长为2 fs。体系先在恒体积恒温(NVT)条件下,再在恒压恒温(NPT)条件下进行能量最小化与平衡。生产模拟阶段采用Nose−Hoover恒温器和Parrinello−Rahman压强耦合,将温度维持在310.15 K,压力维持在1 atm。
为了估算PIEZO2开放孔道的电导,在z方向施加±0.6 V的恒定外加电场,同时在整个模拟过程中保持侧向压力为−3 bar。每个电压条件下进行3个重复的100 ns模拟。通过统计离子从胞外向胞内以及反向穿过孔道的数量来计算离子电流(扩展数据表2)。
4.7 孔道水化与孔内脂质的分析
孔道内粒子数量(扩展数据图3b)是在一个定义的圆柱区域内计算得到的。该圆柱的半径由V2750残基的Cα原子到其几何中心的平均距离确定,高度沿z轴方向在该中心上下各延伸1.5 nm(示意见补充图4)。为了计算脂质头基、脂质尾部、水分子及蛋白在孔道中心周围的径向分布函数(RDF)(图3b及扩展数据图4),将三个V2750的Cα原子的几何中心定义为孔道中心。RDF以该点为圆柱轴心进行计算,其中圆柱半径为6 nm,高度分别设为脂质7 nm、水5 nm、蛋白3 nm。对于每个目标粒子,计算其到圆柱轴线的径向距离,即粒子到垂直轴的垂直距离。径向分布函数
4.8 电生理实验
将编码野生型小鼠PIEZO1或PIEZO2的质粒(1 µg)转染至12孔板中约80%汇合的HEK293TPIEZO1KO细胞中。转染后次日将细胞重新铺板于直径12 mm的玻璃盖玻片上,并在转染后36−48小时内进行实验。膜片钳电极由G150F硼硅玻璃毛细管制备,使用P-97拉制仪拉制,并通过MF2微锻仪进行热抛光。
在图1所示的贴附膜压力钳实验中,电极内液为140 mM KCl、10 mM HEPES、10 mM TEA和2 mM EGTA(pH 7.4),浴液使用含150 mM NaCl的商业HBSS溶液。用于孔径测量的单价阳离子溶液由10 mM HEPES、10 mM EGTA以及150 mM盐溶液组成(包括NaCl、甲胺氯化物、二甲胺氯化物、三甲胺氯化物或四甲胺氯化物),pH通过NMDGOH或HCl调节至7.4。装填这些溶液后电极电阻为2−3 MΩ。切除膜片在HBSS溶液中形成,并通过加压自动灌流系统依次切换不同溶液进行实验。
甲基化胺的相对通透性(PMA)通过修正的Goldman−Hodgkin−Katz方程计算,该方程考虑了PIEZO1和PIEZO2对氯离子具有非零通透性的情况:
其中,
孔道半径基于排除体积理论通过以下公式估算:
其中,
其中,
压力脉冲通过高速压力钳装置经膜片电极施加。压力诱导电流在电压钳模式下以10 kHz采样频率记录,使用Axopatch 200B放大器采集并通过Digidata 1550B进行数字化处理。在每次测量之间,膜片在+5 mmHg和0 mV条件下保持20 s以确保通道完全从失活状态恢复。浴液接地采用Ag/AgCl(3 M KCl)参比电极。在未转染细胞的切除膜片中测得的液接电位在所有测试溶液条件下均小于2 mV,因此在I−V曲线中未进行校正。
单通道记录按照既有方法进行。通过施加一个10 ms、−90 mmHg的压力脉冲估算膜片中通道数量,随后在0 mmHg条件下保持20 s,再在−40 mmHg条件下记录20 s。单通道数据以10 kHz采样并在100 Hz下滤波以分辨次导通状态。全点直方图分析及高斯拟合使用相应软件完成。